
1.20: Fotosíntesis
- Page ID
- 56437

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Las plantas crecen de la nada. Mientras que una pequeña porción de su masa (peso seco) proviene del suelo, aproximadamente 98% proviene de la atmósfera. Una porción típica de papas es de 6 oz = 170 gramos. Dado que las papas son aproximadamente 80% de agua, una porción de papas es de aproximadamente 34 gramos de peso seco. Este peso es almidón casi puro que tiene una fórmula química (C 6 H 12 O 6). Los átomos de carbono representan el 40% del peso del almidón (o de cualquier polímero de hexosa o hexosa) de manera que 34 gramos de papa tienen 13.6 gramos de carbono = 1.13 moles de carbono. Esto se ha derivado del aire que es aproximadamente 400 ppm de dióxido de carbono. Para adquirir el carbono necesario para una porción de papas la planta tiene que extraer carbono de aproximadamente 67 mil litros de aire. Esta capacidad fotosintética es aún más notable porque implica una transferencia de la energía de la luz (radiación electromagnética) a la energía química presente en el carbohidrato que se produce. Las hexosas sintetizadas tienen considerablemente más energía que las materias primas (dióxido de carbono y agua) utilizadas para producirlas.
TEMAS
- Descripción general
- Las reacciones dependientes de la luz
- Las reacciones de asimilación del dióxido de carbono
- Restricciones fotosintéticas: el compromiso de fotosíntesis/transpiración
- Fotosíntesis C4
- Fotosíntesis CAM
Descripción general
La fotosíntesis produce carbohidratos. El nombre revela lo que originalmente se consideraron: los carbonos hidratados (es decir, el agua agregada al carbono), pero la investigación ha revelado que una descripción más precisa de los carbohidratos es el dióxido de carbono reducido. Esto se ilustra en la ecuación general para la fotosíntesis:
CO 2 + H 2 O —> (CH 2 O) n + O 2
Se agrega hidrógeno al dióxido de carbono, es decir, se reduce, y el hidrógeno se está eliminando del oxígeno, es decir, se está oxidando. Obsérvese que, como en el caso de la respiración celular, la ecuación general es un resumen del efecto neto de múltiples reacciones que tienen lugar simultáneamente. Específicamente en fotosíntesis, el dióxido de carbono NO reacciona con el agua. En cambio, tanto el agua como el dióxido de carbono se consumen en un grupo de reacciones que finalmente producen oxígeno y carbohidratos.
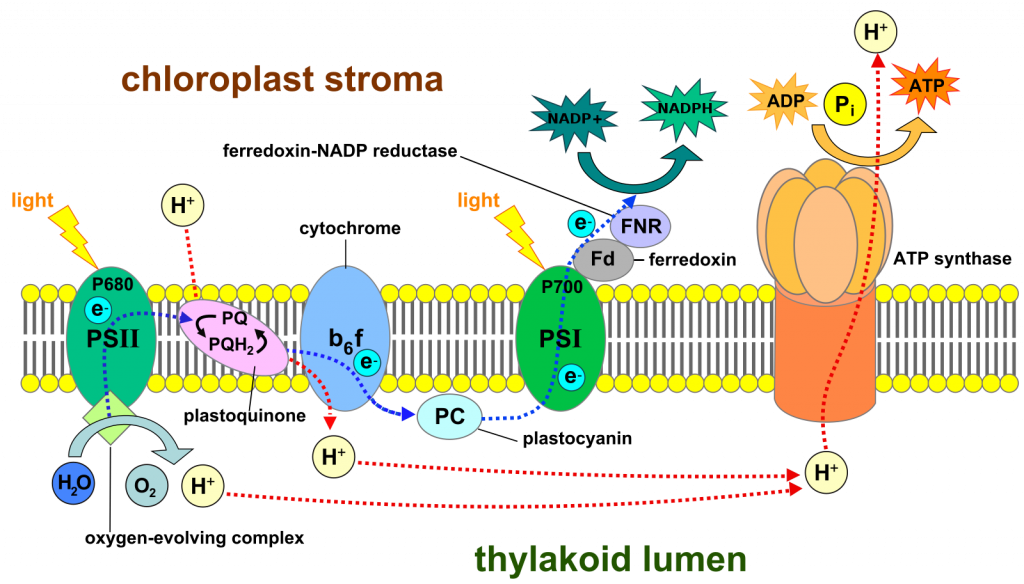
La reacción general tal como está escrita es excepcional porque el oxígeno es un átomo altamente electronegativo, uno que atrae fuertemente a los hidrógenos, mucho más fuertemente que el átomo de carbono al que se transfieren los hidrógenos. Por esta razón, la reacción es 'cuesta arriba' y es poco probable que ocurra, mientras que la reacción en sentido contrario es mucho más probable que ocurra, siendo los carbohidratos oxidados por el oxígeno para producir dióxido de carbono y agua (ver el capítulo anterior). El último capítulo ilustró que la respiración celular (oxidación de carbohidratos) es 'impulsada' por la electronegatividad del oxígeno; esta electronegatividad 'tira' de electrones a través de la membrana mitocondrial interna, uniéndolos finalmente con oxígeno. Por lo tanto, una pregunta significativa sobre la fotosíntesis es qué aleja los electrones del oxígeno de una molécula de agua, ¿qué tiene una 'atracción' más fuerte para los hidrógenos que el oxígeno (es decir, ¿qué es un oxidante más poderoso que el oxígeno)? La luz que interactúa con el pigmento clorofila juega un papel crítico en la generación del oxidante fuerte. También es crítica la estructura del cloroplasto que, al igual que las mitocondrias, consiste en una estructura compleja de membranas que separan los compartimentos acuosos. Las membranas contienen clorofila (Figura 1) así como otros pigmentos y proteínas, organizados de formas muy específicas.

Las reacciones de fotosíntesis dependientes de la luz
El papel de la luz en la fotosíntesis es reorganizar moléculas específicas de clorofila, haciendo que pierdan electrones (oxidándolos) y con ello convirtiéndolos en oxidantes que eliminan electrones de otras moléculas y finalmente forman agua.La luz es una forma de energía y en consecuencia es capaz de cambiar las circunstancias de el material con el que interactúa. En particular, la luz cambia la configuración electrónica de la clorofila, desplazando un electrón de su posición normal a una situación en la que es más probable que escape de la molécula de clorofila, es decir, después de absorber la luz, es más probable que la clorofila se oxide. La oxidación de la clorofila se hace aún más probable porque hay una molécula cercana en la membrana interna del cloroplasto que es capaz de aceptar el electrón. El electrón perdido de la clorofila en última instancia (después de muchos pasos) termina asociado con el carbono de una molécula de dióxido de carbono, formando carbohidratos. La molécula de clorofila oxidada (es decir, a la que le falta un electrón) no es un oxidante lo suficientemente fuerte como para extraer electrones del agua. Pero puede actuar sobre el 'complejo evolutivo de oxígeno', un complejo enzimático que contiene cuatro átomos de manganeso. Una clorofila que carece de un electrón es capaz de oxidar uno de los cuatro átomos de manganeso. Después de que este proceso se repita tres veces más y se oxidan los cuatro átomos de manganeso, la molécula es ahora un oxidante lo suficientemente fuerte como para actuar sobre dos moléculas de agua, eliminando cuatro electrones, uno cada uno de los cuatro hidrógenos, y produciendo cuatro protones (H +) y una molécula de O 2.
El electrón perdido de la clorofila sigue un camino a través de una membrana similar al flujo de electrones a través de la membrana mitocondrial interna en el proceso de 'transporte de electrones' (= fosforilación oxidativa) de la respiración celular, a veces utilizando portadores de electrones similares.
Y, similar a algunos de los pasos en la fosforilación oxidativa, algunas de las transferencias de electrones tienen el efecto de mover protones de un lado de la membrana al otro. Además, la 'división' del agua, realizada por la proteína que contiene manganeso, agrega protones a ese mismo lado de la membrana. La acumulación de protones en un lado de la membrana crea un gradiente electroquímico a través de la membrana. Y, debido al gradiente electroquímico, el ATP se puede sintetizar ya que está en la cadena de transporte de electrones de la respiración celular. Pero en este caso los electrones no están fluyendo al oxígeno. Se mueven primero a un átomo de clorofila diferente, pero solo después de que, como la molécula de clorofila descrita anteriormente, se haya oxidado por la acción de la luz. Cuando esta segunda clorofila recibe un electrón de la cadena de transporte de electrones se convierte de nuevo a su estado normal, y en este estado la clorofila puede absorber una vez más la luz, excitarse y perder un electrón, continuando así el proceso. Tenga en cuenta que hemos identificado dos moléculas distintas de clorofila, las cuales absorben la luz y pierden electrones. Una molécula de clorofila (llamada clorofila 680) obtiene electrones de 'reemplazo' de una proteína que contiene manganeso; la segunda clorofila (llamada clorofila 700) obtiene electrones de reemplazo de la cadena de transporte de electrones. El flujo de electrones descrito hasta ahora es:
agua—>
complejo enzimático de manganeso —>
clorofila 680—>
cadena de transporte de electrones —>
clorofila 700.

¡Pero aún no han terminado su viaje! En última instancia estos electrones estarán reduciendo el dióxido de carbono, pero antes de llegar al carbono de un dióxido de carbono son transferidos a otro intermedio importante, el NADP + (Figura 2), un compuesto muy similar al NAD + que opera en las mitocondrias durante la respiración celular. Al igual que NAD +, NADP + puede aceptar dos electrones y un protón para formar NADPH y puede perder los mismos elementos para reformar NADP +, es decir, es un portador de electrones que (como NADP +) puede oxidar compuestos y (como NADPH) puede reducir compuestos. El NADP + recibe electrones de moléculas portadoras que los reciben de una molécula de clorofila 700 'excitada'. NADPHIS es una molécula relativamente estable y es soluble en agua, a diferencia de muchos de los portadores de electrones involucrados en la fotosíntesis que son solubles solo en los lípidos de la membrana del cloroplasto.
En total, lo que hemos descrito hasta ahora es un flujo de electrones impulsado por la luz del agua al NADP +, formando NADPH y O 2 (Fig 3). El flujo ocurre en la membrana interna del cloroplasto e implica dos etapas donde la energía de la luz es significativa para hacer más probable la transferencia de electrones (es decir, reacciones redox). El flujo de electrones a través de una membrana es capaz de crear un gradiente de protones, como lo hace en la membrana mitocondrial interna. Y, como es el caso en las mitocondrias, este gradiente en protones puede ser utilizado para sintetizar ATP. Este grupo de reacciones, impulsadas por la luz y que crean NADPH y ATP a partir de NADP +, ADP y fosfato inorgánico, se llama 'las reacciones de luz' y se resume como:
H 2 O + NADP + + ADP + iP —> NADPH + ATP + O 2
Las reacciones de asimilación de dióxido de carbono de la fotosíntesis
Los productos de las reacciones de luz, NADPH y ATP, se utilizan para sintetizar carbohidratos a partir del dióxido de carbono, un proceso llamado fijación de dióxido de carbono.
La fijación de carbono implica fundamentalmente el uso del 'poder reductor' del NADPH para reducir el dióxido de carbono, y el proceso se resume en la siguiente ecuación:
NADPH + ATP + CO2—> (CH 2 O) n + NADP + + ADP + iP
Cabe destacar que uno de los aspectos extremadamente importantes de estas reacciones es que regenera los metabolitos necesarios en las reacciones de luz: NADP +, ADP e iP. Dado que los suministros de estos metabolitos son limitados, es crítico que se reciclen. Las reacciones de fijación de carbono se pueden resumir en tres etapas: carboxilación, reducción y regeneración.
Carboxilación
La carboxilación describe la incorporación de dióxido de carbono en una molécula orgánica. Curiosamente, esto se puede lograr sin involucrar a ninguno de los productos de la reacción de luz. La carboxilación ocurre cuando se agrega dióxido de carbono a un metabolito llamado bifosfato de ribulosa (RuBP), un azúcar de cinco carbonos con dos fosfatos, en una reacción catalizada por una enzima llamada ribulosa bisfosfato carboxilasa (rubisco). El compuesto resultante de 6 carbonos se descompone rápidamente en dos moléculas de un compuesto de tres carbonos llamado fosfoglicerato (PGA).
Reducción
Aunque el dióxido de carbono ha sido asimilado, el PGA no es un compuesto muy útil porque está demasiado oxidado. Para ser útil es necesario reducir la PGA. Luego se puede utilizar como molécula precursora para elaborar una variedad de biomoléculas como azúcares, aminoácidos, ácidos nucleicos y muchos otros. Además, el compuesto reducido se puede utilizar para hacer más RUBP y así permitir que se asimile más dióxido de carbono.
La reducción de PGA se logra utilizando NADPH y ATP producidos en las reacciones de luz de la fotosíntesis y produce un azúcar de tres carbonos llamado gliceraldehído-3-fosfato (G3P).
PGA + NADPH + ATP —> G3P + NADP + + ADP + iP
Regeneración de RuBP
Para sostener la fotosíntesis la planta necesita regenerar RUBP, el azúcar de 5 carbonos que se utiliza para adquirir CO 2. Esto ocurre cuando se sintetiza RUBP a partir de G3P. Obviamente, no se puede hacer un azúcar de cinco carbonos con un azúcar de tres carbonos. Podrías hacerlo usando dos moléculas G3P pero habría uno 'fijo' de carbono sobrante. Sin embargo, la síntesis se puede lograr de forma ordenado si se inicia con cinco moléculas de G3P (quince carbonos totales) y se producen tres RUBP' s (también 15 carbonos). Estas reacciones se llaman el ciclo Calvin-Benson y requieren un ATP hecho en las reacciones de luz por cada RUBP producido.
Al mismo tiempo, G3P se puede utilizar para elaborar azúcares de seis carbonos, en particular glucosa y fructosa y a partir de ellos, sacarosa, almidón, celulosa y una amplia variedad de polisacáridos.
Al juntar ambas actividades, si se fijan seis moléculas de dióxido de carbono carboxilando seis RUBP, entonces se pueden producir 12 G3P después de la reducción utilizando 12 NADPH y 12 ATP. Diez moléculas de G3P se pueden usar para regenerar los SixRBP y este proceso requiere seis ATP más. Las dos moléculas restantes de G3P se pueden usar para formar una fructosa o una glucosa. Es así como todas las plantas realizan la fotosíntesis. Cada dióxido de carbono asimilado requiere dos NADPH y tres ATP.
Tenga en cuenta que TODAS las ecuaciones anteriores son resúmenes de múltiples reacciones que ocurren simultáneamente. Hay una gran cantidad de química ocurriendo en los cloroplastos, aunque el efecto neto puede expresarse simplemente como
6 CO 2 + 6 H 2 O —> (C 6 H 1 2 0 6) + 6 O 2
Tenga en cuenta que la ecuación neta no incluye ATP, ADP, iP, NADP +, NADPH, RuBP, PGA o G3P. Todos estos reactivos se producen al mismo ritmo que se consumen. La ecuación neta también oculta el hecho de que en realidad se necesitan 12 H 2 O como reactivos y que 6 H 2 O son productos (el efecto neto es simplemente el consumo de 6 H 2 O). Parte de la elegancia de la fotosíntesis es que a pesar de la miríada de reacciones que tienen lugar, el efecto neto es muy simple. Otro aspecto de su elegancia es que el producto del proceso, el carbohidrato (CH 2 O) n, se puede utilizar para elaborar todos los diversos químicos que produce la planta, no solo los obvios (almidón, celulosa, hemicelulosa, pectinas) sino los menos obvios (aminoácidos, ácidos nucleicos , lípidos, lignina, etc, etc.). Además, los carbohidratos también se utilizan como fuente de energía para alimentar estas reacciones sintéticas: En la respiración celular.los carbohidratos se consumen para producir dióxido de carbono y agua mientras producen ATP y NADH, químicos que se necesitan en muchas de las reacciones sintéticas.
Adquisición de dióxido de carbono, problemas y soluciones
El dióxido de carbono fijado en la fotosíntesis se deriva de la atmósfera. En los últimos 150 años, los niveles de dióxido de carbono atmosférico han aumentado en más de 40% pero siguen siendo muy, muy bajos a 0.04% o 400 ppm (partes por millón; por cada millón de moléculas en el aire solo 400 de ellas son dióxido de carbono). El dióxido de carbono ingresa a la hoja por difusión, pero como la concentración máxima en el exterior es de 400 ppm y, para la mayoría de las plantas, la concentración mínima en el interior es de aproximadamente 100 ppm. Este mínimo es el resultado de que el rubisco no puede carboxilar RUBP si las concentraciones de dióxido de carbono están por debajo de 100ppm, de ahí que el [CO 2] no pueda ir por debajo de 100 ppm. En consecuencia, la fuerza impulsora para la difusión (la diferencia en [CO 2] entre el interior y el exterior de la hoja) es baja y potencialmente está limitando el flujo de dióxido de carbono hacia la hoja y consecuentemente la fotosíntesis. Considerando la ecuación de flujo (Capítulo 3), el flujo de dióxido de carbono hacia la hoja puede incrementarse disminuyendo la resistencia al movimiento, es decir, haciendo que la hoja sea más porosa a la atmósfera abriendo estómagos, los poros regulados en la epidermis foliar. Desafortunadamente para la mayoría de las plantas, esta exposición a la atmósfera resulta en pérdida de agua, denominada transpiración. El agua perdida necesita ser reemplazada por agua del suelo, agua que a veces escasea. Esto lleva a lo que se ha denominado el compromiso de fotosíntesis/transpiración: en la medida en que la planta gana dióxido de carbono para la fotosíntesis, pierde agua en la transpiración.
Las bajas concentraciones de dióxido de carbono en el aire agravan el problema: permitir que más dióxido de carbono ingrese a la hoja puede abrir sus estómagos más completamente pero esto da como resultado la pérdida de más agua. En ambientes húmedos, donde el agua está fácilmente disponible, no hay problema con tener hojas muy porosas (es decir, estómagos abiertos), pero cuando el agua es escasa la planta debe equilibrar la ganancia de carbono con la pérdida de agua. El comportamiento estomático refleja este compromiso: generalmente cierran por la noche, cuando la fotosíntesis es imposible, y en tiempos de sequía, al adquirir agua para reemplazar lo que se ha perdido es difícil.
El dióxido de carbono adicional en la atmósfera en los últimos 200 años probablemente explica la observación de que la tierra es 'más verde' (mayor cobertura foliar) ahora que hace 50 años. Más dióxido de carbono puede actuar como 'fertilizante' y/o permite que las plantas sobrevivan en condiciones de bajo suministro de agua.
Si bien hay una serie de características que permiten que algunas plantas sobrevivan en condiciones secas, hay una que involucra directamente las reacciones fotosintéticas discutidas anteriormente. Esta vía fotosintética modificada, llamada vía C4, permite que algunas plantas adquieran dióxido de carbono al tiempo que pierden menos agua de lo que haría una planta normal. Las plantas C4 concentran dióxido de carbono en una porción relativamente pequeña de la hoja, llamadas células de vaina de haz. La vía utiliza dos carboxilaciones. La primera ocurre en las células del mesófilo foliar y utiliza una enzima, PEP carboxilasa, que agrega un carbono a un compuesto de tres carbonos, fosfoenolpiruvato (PEP). PEP carboxilasecan opera a niveles de dióxido de carbono de hasta alrededor de 10 ppm, aproximadamente 1/10th de la concentración necesaria para que el rubisco opere. El compuesto de cuatro carbonos producido por PEP carboxilasa se transporta vía plasmodesmata a una vaina de células agrandadas (Figura 4) que rodea las hebras vasculares de la hoja. Aquí el compuesto de cuatro carbonos es descarboxilado, liberando dióxido de carbono que posteriormente se fija en fotosíntesis normal usando rubisco. Los tres fragmentos de carbono restantes se transportan de regreso a las células mesófilas. El uso de este sistema de dos carboxilaciones permite a las plantas producir un ambiente (las células de la vaina del haz) donde hay una mayor concentración de dióxido de carbono. Es sólo en esta ubicación donde está presente rubisco. Debido al mecanismo de concentración de CO 2, el [CO 2] puede ser mayor a 100 ppm en las células de vaina del haz mientras que el [CO 2] del aire dentro de la hoja está cerca es menor de 10 ppm. Debido a que la concentración de CO 2 es sustancialmente menor que en una hoja normal, puede haber una mayor fuerza impulsora para la difusión en la hoja. Esto permite que la resistencia sea mayor (estoma más cerrado) al tiempo que se logra la misma cantidad de flujo de dióxido de carbono (fotosíntesis) que una planta que no utilizó esta vía. Así, la hoja puede funcionar fotosintéticamente a la vez que es mucho menos porosa al dióxido de carbono, perdiendo así menos agua en la transpiración.

Las plantas que utilizan esta ruta de carboxilación dual se denominan “plantas C4” porque la primera carboxilación produce compuestos de cuatro carbonos opuestos al patrón en la mayoría de las plantas, llamadas plantas C3, donde la carboxilación produce un compuesto de tres carbonos. La ruta C4 se describe a continuación:
- en las células mesófilas de la lea f un compuesto de tres carbonos, PEP, es carboxilado por PEP carboxilasa, para formar un compuesto de cuatro carbonos
- el compuesto de cuatro carbonos se transporta a las células de la vaina del haz que rodean los haces vasculares (las venas de la hoja)
- el compuesto de cuatro carbonos está descarboxilado, liberando CO 2 y piruvato, un compuesto de tres carbonos
- el piruvato es transportado de vuelta a las células mesófilas donde se convierte en PEP en un proceso que requiere energía en forma de NADPH y ATP
- el CO 2 que se liberó en la vaina del haz se fija utilizando rubisco en la vía normal
Tenga en cuenta que C4 requiere de toda la maquinaria y reacciones de la fotosíntesis C3, es solo que hay un conjunto adicional de pasos previos al patrón C3. La fotosíntesis C4 es menos eficiente que la fotosíntesis C3 porque requiere más energía ATP. Sin embargo, es más eficiente en términos de uso del agua y esto es de mayor importancia en regiones más secas. Además, por razones en las que no vamos a entrar, la fotosíntesis C4 se ve favorecida a temperaturas más altas.

Hay un segundo grupo de plantas que utilizan la vía dual de carboxilación pero de manera modificada. Se llaman plantas CAM. CAM se refiere al metabolismo del ácido crassuláceo porque el patrón de comportamiento mostrado por este grupo fue descubierto por primera vez en plantas suculentas de la familia Crassulaceae. Las plantas CAM tienen varias peculiaridades: suelen ser suculentas, es decir, tienen hojas gruesas y carnosas (Figura 5), o no tienen hojas y un tallo grueso y carnoso (por ejemplo, cactus). Por lo general, muestran un marcado patrón diario de acidez tisular con la acidez más alta al amanecer y acidez decreciente durante las horas de luz y acidez creciente durante la noche. Lo más peculiar es que abren sus estómagos por la noche, no durante el día mientras que la mayoría de las plantas solo abren sus estómagos durante el día cuando pueden fotosintetizar.
Básicamente, estas plantas operan igual que las plantas C4 pero en lugar de tener una separación espacial de las dos carboxilaciones como tienen las plantas C4 (mesófila vs vaina de haz), tienen una separación temporal (diurna vs noche). La carboxilación inicial ocurre por la noche cuando los estómagos están abiertos. El compuesto de cuatro carbonos es un ácido y hace que aumente la acidez del tejido. Durante el día se cierran los estómagos, el dióxido de carbono es proporcionado por la descarboxilación de los cuatro ácidos de carbono y la acidez del tejido disminuye. El dióxido de carbono liberado se vuelve a fijar vía rubisco para formar carbohidratos.
¿Estas plantas fotosintetizan de noche o de día? Depende de cómo se quiera definir la fotosíntesis: la adquisición de carbono es de noche pero la síntesis de azúcar es durante el día. La fotosíntesis CAM está asociada, pero no obligatoriamente, con suculencia. Las plantas CAM se encuentran generalmente en hábitats que son secos, ya sea climáticamente, por ejemplo, desiertos, o por microhábitat, por ejemplo, epífitas, plantas que no están enraizadas en el suelo pero que crecen en otras plantas. Las pifitas E suelen estar expuestas a la sequía debido a su falta de conexión con el suelo. Los cactus y otras suculentas sin hojas son comúnmente plantas CAM. Las tasas fotosintéticas de CAM son muy bajas y las tasas de crecimiento de las plantas CAM también son bajas. La asociación de suculencia y CAM probablemente refleja el hecho de que las plantas en hábitats áridos a menudo desarrollan suculencia para almacenar agua y que la suculencia es más propicia para la CAM porque los tejidos suculentos pueden almacenar más carbono como el ácido de cuatro carbonos.
Los patrones de fotosíntesis CAM y C4 revelan que estas vías han evolucionado varias veces, es decir, las vías dicen poco sobre la filogenia. C4, C3 y CAM se mezclan dentro de géneros, familias y órdenes. Al parecer, es relativamente 'fácil' para C4 y CAM evolucionar.