
1.10: Vías genéticas
- Page ID
- 56652

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)- Comprender cómo múltiples genes a menudo funcionan juntos para controlar un rasgo
- Comprender el concepto de una vía biosintética
- Comprender el concepto de una vía regulatoria
- Comprender el concepto de una red
- Entender cómo las mutaciones en diferentes genes de una vía pueden causar el mismo fenotipo
- Entender cómo las mutaciones en diferentes genes de una vía pueden causar diferentes fenotipos
Introducción
Los genes no funcionan aisladamente, sino que conjuntos de genes actúan en concierto para realizar funciones biológicas. Cuando diferentes genes funcionan en diferentes etapas secuenciales de un proceso biológico, esto se conoce como una vía genética. Quizás el tipo de vía más intuitiva conceptualmente es una vía biosintética, donde una molécula precursora se modifica químicamente a través de una serie de etapas intermedias catalizadas enzimáticamente para producir un producto bioactivo. Dado que las enzimas son (proteínas), los genes que codifican estas enzimas se consideran una vía genética. En una vía reguladora, algún tipo de estímulo conduce a un cambio en la expresión o actividad de un determinado producto génico, que a su vez actúa para alterar la expresión o actividad de otro producto o productos génicos, que a su vez podrían regular otro nivel de actividad.
En última instancia, estos cambios reguladores en la expresión o actividad conducen a una respuesta al estímulo, generalmente involucrando cambios en la expresión génica. Nuevamente, los genes que codifican los diversos tipos de moléculas reguladoras, principalmente proteínas, constituyen una vía genética. Las vías suelen ser objetivos en la manipulación biotecnológica de los rasgos, lo que requiere una comprensión de cómo se comportan las vías. También hay que señalar que muchos sistemas biosintéticos y reguladores se describen mejor como redes debido a que a menudo, las vías se ramifican o convergen e interactúan con otros sistemas. Sin embargo, por simplicidad tales redes complejas no serán discutidas aquí.
Vías biosintéticas
Los genes codifican enzimas que catalizan etapas en la síntesis de un compuesto. Una ruta biosintética describe realmente un proceso de conversión de un sustrato precursor en un producto. Además del sustrato precursor y producto, incluye las enzimas, las reacciones químicas catalizadas por las enzimas y todos los compuestos intermedios. Una vía consiste en una serie de etapas donde una molécula precursora actúa como sustrato para una enzima, la cual cataliza una reacción química para producir un producto, que luego sirve como sustrato para una etapa posterior.

Ya hemos discutido el gen, una hipótesis enzimática que fue un avance importante en el descubrimiento de cómo los genes codifican proteínas. Este análisis también proporciona un buen ejemplo del concepto de una vía biosintética. Esta hipótesis se descubrió mediante el análisis de mutantes de neuroesporas auxotróficas que eran defectuosos en la producción de aminoácidos. Es decir, las vías biosintéticas que producen aminoácidos particulares fueron alteradas por mutaciones. El texto discute el análisis de la vía de la arginina. Los mutantes que interrumpen esta vía no pueden crecer en un medio mínimo que carece de arginina, pero pueden crecer si se suministra arginina en el medio. La misma vía fundamental también produce arginina en las plantas.
Las mutaciones que interrumpen la función de enzimas particulares en la ruta bloquean la progresión de la ruta en ese paso correspondiente. Por ejemplo, una mutación en el gen argF alteraría la enzima ornitina carbamoiltransferasa (OTC). Esto bloquearía el paso 6 en la vía, la conversión de L-ornitina a L-citrulina. Como resultado, el compuesto intermedio L-Ornitina probablemente se acumulará, mientras que todos los compuestos que se presentan más adelante en la ruta (es decir, L-Citrulina, L-argininosuccinato y L-arginina) serán deficientes.
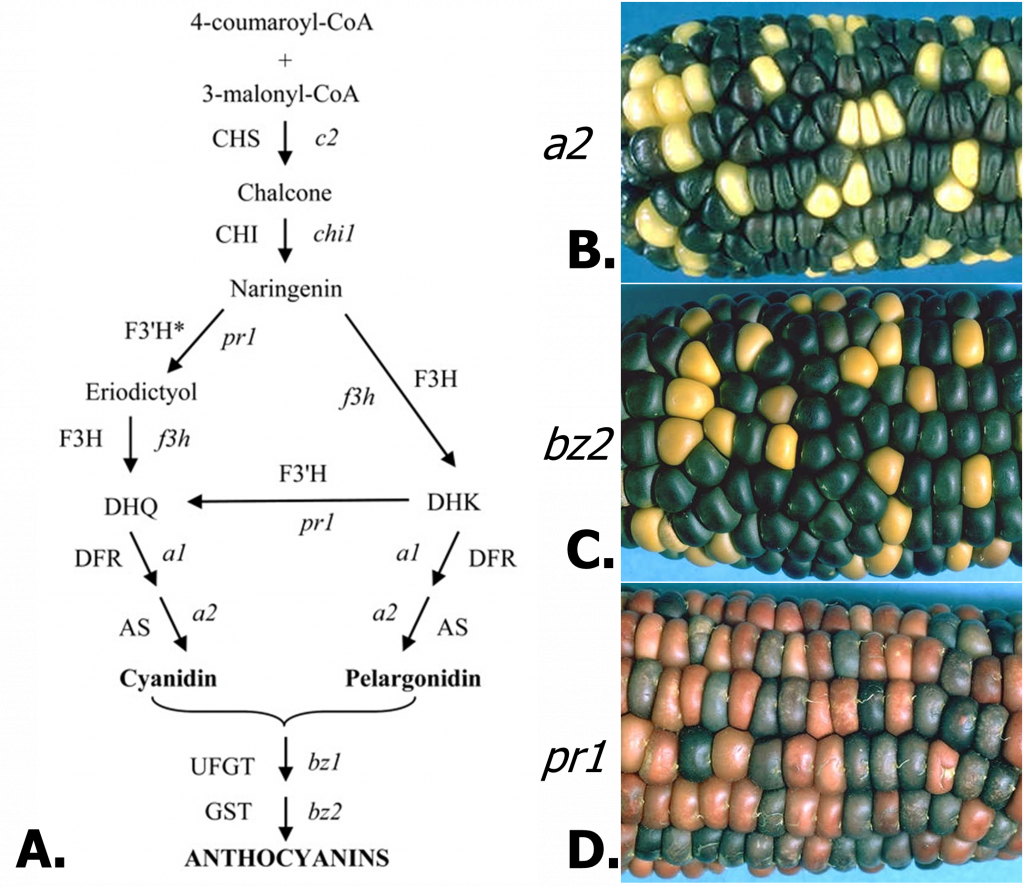
La vía de las antocianinas
La vía de antocianinas es una vía un poco más compleja, aunque sigue siendo simple en lo que respecta a estas cosas. Es de gran interés para los horticultores porque los pigmentos de antocianina son responsables de muchos colores vegetales, particularmente en flores y frutos (piense en el vino tinto). Los pigmentos de antocianina varían de naranja a rojo, púrpura y azul. Además de los atractivos colores que proporcionan las antocianinas, también existe un considerable interés reciente en las antocianinas por su valor dietético, actuando como fuertes antioxidantes y posiblemente agentes anticancerígenos.
Efectos de mutaciones en diferentes etapas en la biosíntesis de antocianinas.
Por supuesto, cada enzima en la ruta de antocianinas está codificada por un gen. Como tal, las mutaciones pueden generar variación o interrumpir la ruta. La ruta de antocianina del maíz y ejemplos de mutantes se muestran en la Figura 2. El mutante a2 provoca una deficiencia en la enzima antocianidina sintasa, bloqueando la producción de cualquier compuesto pigmentado. Los mutantes a1 y c2 son igualmente incoloros. El mutante bz2 es deficiente en una enzima relacionada con la glutatión-S-transferasa, la cual se requiere para el transporte de antocianidinas a la vacuola, dando como resultado el pigmento de color bronceado debido al ambiente más alcalino del citosol. El mutante bz1 (no mostrado) produce un fenotipo similar. La mutación del gen pr1 elimina solo una clase de antocianinas. El gen pr1 codifica flavanona 3′-hidroxilasa, requerida para la producción de cianidina, la cual genera antocianina púrpura. En el mutante, solo se sintetiza pelargonidina, produciendo un pigmento rojo.
Vías regulatorias
Una vía reguladora se refiere a una situación en la que un gen o producto génico controla la expresión o actividad de otro gen o producto génico, que a su vez también puede realizar una función reguladora. Como discutimos, la función génica puede regularse en muchos puntos, desde la regulación transcripcional hasta la regulación postraduccional. Examinaremos un par de ejemplos específicos de vías reguladoras, pero primero consideremos algunas de las características generales de las vías reguladoras. Comprender la lógica de las vías reguladoras será importante más adelante cuando busquemos utilizar la biotecnología para manipular rasgos alterando su regulación.
Una manera simplista pero útil de ayudar a comprender la lógica de las vías regulatorias es considerarlas como una serie de interruptores con estados de encendido y apagado.
Reguladores positivos vs negativos
Una característica clave de las vías reguladoras es que los reguladores pueden funcionar positiva o negativamente. Un regulador positivo es aquel que funciona para activar el siguiente componente en la vía, mientras que un regulador negativo inhibiría al siguiente componente. Por ejemplo, un factor de transcripción que activara la transcripción de un gen se consideraría un regulador positivo, mientras que uno que reprimiera la transcripción de un gen sería un regulador negativo. Otro tipo común de regulador es una proteína quinasa. Las proteínas quinasas son proteínas que agregan un grupo fosfato a otra proteína. La fosforilación es un tipo común de modificación postraduccional que puede regular la actividad de las proteínas; a veces la proteína fosforilada se activa y a veces se reprime.
Un paso regulatorio positivo se representa con una flecha. Por ejemplo, “la proteína A activa la proteína B” se representaría como:
\[\text{A} \to \text{B}\]
Un paso regulatorio negativo se representa con una barra. “La proteína C reprime la proteína D” estaría representada como:
\[\text{C} \dashv \text{D}\]
Estados activos vs. estados inactivos de reguladores
Un segundo punto a considerar en la lógica de las vías reguladoras es el estado de actividad de cada componente. Una manera útil de pensar sobre esto es que cada componente puede tener un estado “encendido” o “apagado”. Por supuesto, en realidad, muchas proteínas pueden tener niveles intermedios de actividad pero en aras de la simplicidad, solo consideraremos los estados de encendido y apagado.
El efecto de que un regulador esté en estado encendido o apagado depende de si es un regulador positivo o negativo.
Si un regulador positivo está en el estado activado, funcionará para activar el siguiente factor. En el ejemplo de , si A está en el estado activado o activo, funcionará para cambiar B al estado activado o activo.
, si A está en el estado activado o activo, funcionará para cambiar B al estado activado o activo.
Si un regulador negativo está en el estado encendido, funcionará para cambiar el siguiente factor al estado apagado o inactivo. Porque , si C está en el estado activado o activo, cambiará D al estado apagado o inactivo.
, si C está en el estado activado o activo, cambiará D al estado apagado o inactivo.
Las vías reguladoras a veces contienen muchos, a veces pocos, pasos. También pueden contener una mezcla de reguladores positivos y negativos. Exploremos algunos ejemplos hipotéticos para ver cómo encajan las piezas para regular las respuestas de las plantas a los estímulos.
Cuando se representa un camino, se muestran todos los pasos. En aras de la simplicidad, se supone que el estado de actividad predeterminado de cualquier componente dado es tal que el paso anterior funciona para cambiarlo. Comenzaremos con un ejemplo sencillo que contiene varios pasos reguladores positivos que activan una actividad celular, la Actividad 1, en respuesta al Estímulo 1. En la vía que se muestra a continuación, el Estímulo 1 activa A. Por lo tanto, suponemos que a falta de este estímulo, A se encuentra en estado inactivo o apagado. Dado que A está inactivo, no está funcionando para activar B, que por lo tanto también se encuentra en el estado inactivo, y de igual manera para C. En última instancia, la Actividad 1 no ocurre en ausencia del Estímulo 1 sino que ocurre en respuesta al estímulo.
Estímulo 1 → A → B → C → Actividad 1
Podemos usar códigos de colores para ayudar a visualizar esto. El gris claro representa el estado OFF y el azul representa el estado ON.
Sin Estímulos 1 → A → B → C → Actividad 1 (DESACTIVADO)
Estímulo 1 → A → B → C → Actividad 1 (ON)
O podemos hacer una mesa para ayudar a realizar un seguimiento de los estados de cada componente en presencia o ausencia de estímulo.
| Estímulo 1 → | A → | B → | C → | Actividad 1 |
| NO | OFF | OFF | OFF | NO |
| SI | EN | EN | EN | SI |
Ahora veamos otro ejemplo que contiene reguladores tanto positivos como negativos.
Estímulo 2 → D → E F → Actividad 2
F → Actividad 2
En este caso, los componentes D y E estarían en los estados inactivos en ausencia de estímulo, porque no habrían sido activados. Pero E funciona como regulador negativo de F. Dado que E está inactivo, no está funcionando para inhibir F, que luego permanece en estado activo promover la Actividad celular 2. Cuando el Estímulo 2 está presente, D y E se activan, y E funciona para inactivar F. Dado que F está apagada, la Actividad 2 no ocurre. Así, la respuesta neta al Estímulo 2 es la represión de la Actividad 2.
\[\dashv\]
\[\dashv\]
O
| Estímulo 2 → | D → | E |
F → | Actividad 2 |
| NO | OFF | OFF | EN | SI |
| SI | EN | EN | OFF | NO |
Ahora que hemos considerado las vías regulatorias desde una perspectiva hipotética, consideremos un par de vías reales. Como se mencionó, las vías regulatorias pueden tomar muchas formas. Pueden consistir en una serie de factores de transcripción que regulan la expresión de los genes de los demás, resultando en última instancia en la regulación de genes que afectan alguna respuesta biológica. Aquí, veremos una vía de transducción de señales que consiste en una serie de modificaciones postraduccionales que ocurren en respuesta a un estímulo hormonal y culminan en cambios de expresión génica para efectuar una respuesta.
Vía de transducción de señales para la señalización hormonal
Los brasinoesteroides (BR) son una clase de hormonas esteroides vegetales que controlan muchos aspectos de la fisiología vegetal. Son más destacados por su papel en la promoción del crecimiento de las plantas y los mutantes deficientes en la biosíntesis de BR o señalización muestran enanismo. Los BR también promueven una variedad de otras respuestas, incluyendo el rendimiento y el aumento de la resistencia al estrés.
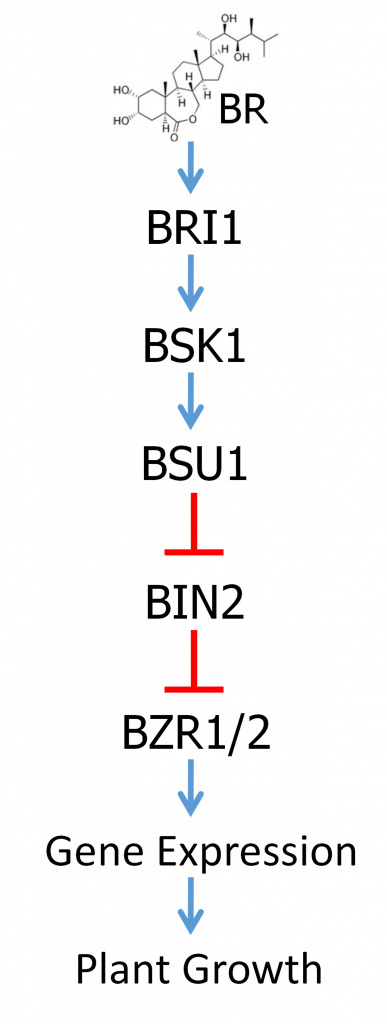
Las hormonas BR, de las cuales la brasinólida (BL) es considerada la más activa, son reconocidas por un receptor llamado BRI1. BRI1 es un tipo de receptor conocido como receptor quinasa, que abarca la membrana celular. En el exterior de la célula hay un dominio receptor que reconoce específicamente y se une a BR. También hay un dominio que abarca la membrana y luego un dominio de proteína quinasa dentro de la célula. La unión a BR fuera de la célula activa entonces el dominio de proteína quinasa dentro de la célula. Las proteínas quinasas son enzimas que funcionan para agregar grupos fosfato a otras proteínas. Como se discutió anteriormente, la fosforilación puede alterar la actividad de una proteína. La activación de la quinasa receptora BRI1 resulta entonces en la activación de una vía de señalización que finalmente regula la actividad de los factores de transcripción dentro del núcleo, cambiando la expresión de genes que regulan el crecimiento y la resistencia al estrés.
La vía de transducción de señales BR está bien estudiada y se resume en la Figura 3. No se preocupe por memorizar los nombres de todos estos factores, sino que solo concéntrese en comprender la lógica regulatoria. Las relaciones regulatorias globales de la ruta se representan en la Figura 3. El objetivo principal de la regulación es un par de factores de transcripción estrechamente relacionados llamados BZR1 y BZR2. BZR1/2 están regulados negativamente por una proteína llamada BIN2 en ausencia de BR. BIN2 es una proteína quinasa que fosforila BZR1/2, lo que resulta en la exclusión de BZR1/2 del núcleo y su degradación proteolítica. En ausencia de BR, BIN2 contiene un grupo fosfato que se requiere para su actividad.
La unión de BR al dominio del receptor BRI1 fuera de la célula da como resultado la activación del dominio quinasa dentro de la célula. BRI1 activado luego fosforila una proteína llamada BSK. La BSK fosforilada se une a otra proteína llamada BSU1, que es una proteína fosfatasa. Las proteínas fosfatasas son enzimas que eliminan los grupos fosfato de otras proteínas. La unión de BSK activa BSU1 que luego cataliza la eliminación del fosfato en BIN2, inactivando así BIN2. Así, BSU1 es un regulador negativo de BIN2. Cuando se inactiva BIN2, eso permite que BZR1/2 se acumule, ingrese al núcleo y regule la expresión de genes que afectan la respuesta BR, incluyendo promover el crecimiento y rendimiento de las plantas.
Efectos de la mutación en diferentes etapas
Las mutaciones pueden afectar las vías reguladoras de múltiples maneras. La mayoría de las mutaciones son mutaciones de pérdida de función donde la función del gen o producto génico está parcial o completamente alterada. Considerando la vía de señalización BR, los fenotipos de mutación diferirán dependiendo de si la mutación afecta a un regulador positivo o negativo de la vía. Una mutación de pérdida de función en el gen bri1 bloqueará la percepción de la hormona y por lo tanto la vía no se activará; se producirá un fenotipo enano. Por otro lado, una pérdida de función en el gen bin2 liberará BZR1/2 de la regulación. El resultado será una actividad constante de BZR1/2 independientemente de si la hormona BR está presente. Se podría esperar que esto genere plantas gigantes, pero de hecho los efectos de la desregulación de las respuestas hormonales son más complejos, llamémoslo crecimiento no regulado.
Las mutaciones también pueden ser ganancia de función, donde el producto génico asume una forma que está constantemente en estado activo. Tales mutaciones son típicamente dominantes y son mucho menos comunes que la pérdida de función. Nuevamente, el resultado de una mutación de ganancia de función en la vía de señalización BR variará dependiendo de si afecta a un regulador positivo o negativo de la vía. Una mutación de ganancia de función en un regulador positivo causaría respuestas hormonales desreguladas, mientras que tal mutación en un regulador negativo cerraría permanentemente la vía y causaría enanismo.
Acción génica complementaria y epistasis genética en el contexto de vías
Considerando que los procesos biológicos están controlados por múltiples genes que actúan juntos, varios principios genéticos tienen más sentido. Primero, queda claro cómo las mutaciones en diferentes genes pueden producir los mismos efectos fenotípicos. Dado que a menudo se requieren múltiples genes para un solo proceso biológico, la interrupción de diferentes genes en el proceso podría tener el mismo efecto neto. De esto se desprende el concepto de acción génica complementaria. Las plantas mutantes con el mismo fenotipo se cruzan entre sí y la descendencia es de aspecto normal. Las mutaciones en las plantas parentales probablemente afectaron a diferentes genes en la misma vía. Finalmente, el concepto de epistasis se puede entender mejor en el contexto de las vías. Tenga en cuenta que el término epistasis es utilizado de manera diferente por diferentes genetistas. En genética cuantitativa, la epistasis se refiere a cualquier interacción génica. Aquí, el término epistasis se refiere a un tipo específico de interacción génica donde la acción de un gen está enmascarada por la acción de otro gen. En otras palabras, si se combinan dos mutaciones en un individuo, solo uno de los fenotipos es aparente. Se dice que el mutante cuyo fenotipo es aparente es epistático al que está enmascarado.
Acción génica complementaria en una vía reguladora
El concepto de acción génica complementaria no es diferente en una vía reguladora que en una vía biosintética. Se esperaría que las mutaciones en diferentes genes que tienen papeles similares en la ruta (por ejemplo, son ambos reguladores positivos de la ruta) causen fenotipos mutantes similares. Si dichos mutantes en diferentes genes se cruzaran entre sí, la progenie F1 sería normal porque ambos genes serían heterocigotos y por lo tanto estaría presente una copia funcional de ambos factores.
Epistasis en una vía reguladora
La epistasis puede ser bastante compleja, especialmente en las vías reguladoras. A diferencia de la acción génica complementaria, la epistasis no se limita a mutaciones recesivas de pérdida de función. La situación se complica aún más por la presencia de reguladores positivos y negativos en una vía. A diferencia de una vía biosintética, la regla general para una vía reguladora es que el gen más “aguas abajo” es epistático al más “aguas arriba” uno. Volvamos de nuevo a la vía de señalización BR y veamos un par de ejemplos para entender por qué.
Primero considere las mutaciones de pérdida de función en los genes bri1 y bin2, que en mutantes individuales causan un fenotipo enano y un fenotipo de crecimiento no regulado, respectivamente. Si las dos mutaciones se combinan en una planta doble mutante, ¿qué esperamos? Las mutaciones bri1 interrumpen la función del receptor requerida para iniciar la señalización a través de la ruta. Sin embargo, BIN2 funciona inhibiendo la actividad de la vía reprimiendo la función de BZR1/2. Si BIN2 se vuelve no funcional, entonces BZR1/2 estará activo y promoverá la respuesta de crecimiento. Esto será cierto incluso si los pasos anteriores están bloqueados por la mutación bri1. Entonces el doble mutante mostraría el fenotipo de crecimiento no regulado bin2 haciendo bin2 epistático a bri1.
¿Qué pasa con una mutación de ganancia de función en BZR1 o BZR2 combinada con una mutación de pérdida de función en bri1? Una mutación de ganancia de función volvería a BZR1/2 constitutivamente activo, independientemente del estado de actividad de BIN2, dando como resultado el fenotipo de crecimiento no regulado. Dado que la actividad de BIN2 sería irrelevante, las actividades upstream también serían irrelevantes. Así, el doble mutante mostraría crecimiento no regulado y BZR1/2 sería epistático a bri1.
Vimos que los genes no actúan solos sino que funcionan en concierto con otros genes para realizar diversas funciones biológicas como la biosíntesis de un compuesto o la regulación de una respuesta a un estímulo. Cuando los genes controlan una serie secuencial de pasos, eso se llama vía. Pero las vías típicamente se ramifican e intersectan con otras vías para formar redes. Las mutaciones afectan la actividad de las vías de varias maneras, dependiendo de la naturaleza de la mutación (pérdida vs ganancia de función) y la función del producto génico (efector positivo vs negativo).
Cuando las mutaciones afectan diferentes etapas en una ruta, a menudo pueden tener un efecto general similar en la ruta y, por lo tanto, conducir a consecuencias fenotípicas similares. Tales mutaciones se complementarán entre sí aunque sus fenotipos sean idénticos. También es posible que las mutaciones en diferentes etapas de una vía puedan causar diferentes fenotipos. Cuando tales mutaciones se combinan en un individuo doble mutante, el fenotipo de uno a menudo está enmascarado por el fenotipo de la otra mutación epistática. Cuando las vías son lineales, la mutación “aguas arriba” es generalmente epistática en las vías biosintéticas y en las vías reguladoras es generalmente la mutación “aguas abajo” la que es epistática. En vías o redes ramificadas, es más difícil generalizar.
La acción génica complementaria es importante para los genetistas que intentan identificar todos los pasos en una ruta. Esta es la base de la prueba clásica de complementación utilizada para determinar si las mutaciones son alélicas o afectan a genes independientes. La epistasis también es útil para genetistas que quieran entender si mutaciones con diferentes fenotipos afectan la misma vía, y si es así, determinar el orden de acción génica.
Cuando la biotecnología se utiliza para manipular un rasgo, es realmente la actividad de una vía o red la que se está atacando. Los mismos principios genéticos descritos en esta sección se utilizan para diseñar estrategias biotecnológicas que alteren las actividades de la vía a fin de producir el resultado deseado sobre el rasgo de interés.
Para responder a las siguientes preguntas, refiérase a las vías ilustradas en la Figura 3. Se etiquetan de manera ligeramente diferente pero a1 codifica DFR, que cataliza la producción de leucoantocianidinas y a2 codifica AS o LDOX/ANS, que posteriormente cataliza la producción de antocianidinas. Supongamos que los granos de maíz normales de tipo silvestre son de color morado/negro profundo como en la Figura 3.
- ¿Qué pronosticarías que pasa con los niveles de leucoantocianidinas en un mutante a2 en comparación con lo normal?
- sin cambios
- aumentar
- disminuir
- no puede predecir
Respuesta: b o d.
La expectativa sería b, que las leucoantocianidinas aumentarían. Debido a que la vía está bloqueada en la etapa posterior, las leucoantocianidinas no pueden convertirse en antocianidinas. Pero como todas las enzimas precedentes siguen siendo funcionales, se esperaría que esas reacciones continuaran dando como resultado la acumulación de leucoantocianidinas.
d también es una respuesta correcta. Las vías biosintéticas no siempre se comportan como se esperaba. Por ejemplo, podría haber inhibición por retroalimentación de una o más enzimas por compuestos intermedios. En pocas palabras, no lo sabremos con certeza hasta que los niveles se midan experimentalmente.
- ¿Qué pronosticarías que pasa con los niveles de pelargonidina en un mutante pr1 en comparación con lo normal?
- sin cambios
- aumentar
- disminuir
- no puede predecir
Respuesta: b o d.
Nuevamente, la expectativa sería b, que la pelargonidina aumentaría. Normalmente, alguna fracción de los compuestos intermedios se canaliza a través de la rama de la ruta que conduce al cianuración. Debido a que la vía que conduce a la cianidina está bloqueada, se podría esperar que todos los intermedios se conviertan en pelargonidina, provocando una mayor acumulación.
d también es una respuesta correcta. Como se discutió en el problema 1, las vías biosintéticas no siempre se comportan como se esperaba. La inhibición de retroalimentación podría ser un factor o puede ser posible que la capacidad enzimática de la rama de pelargonidina de la ruta ya esté saturada. Para aumentar el flujo a través de la vía, podría requerirse aumentar los niveles o actividades de una o más enzimas limitantes de velocidad. Nuevamente, no lo sabremos con certeza hasta que los niveles se midan experimentalmente.
Predecir los resultados de las siguientes mutaciones en la salida de señalización, o la respuesta de la planta para la vía de señalización BR.
- Una mutación de ganancia de función en el gen BZR2
- aumento de la producción
- sin cambios
- disminución de la producción
- no puedo decir
Respuesta: a. Dado que BZR2 es un regulador positivo de la señalización de BR, se esperaría un aumento de la salida de la vía de una mutación de ganancia de función.
- Una mutación de pérdida de función en el gen bsu1. (¡Estudia el camino y piensa en éste!)
- aumento de la producción
- sin cambios
- disminución de la producción
- no puedo decir
Respuesta: c. La producción de la vía disminuiría provocando una disminución del crecimiento de las plantas. A pesar de que BSU1 es un regulador negativo, su objetivo es BIN2 que es otro regulador negativo. Sin la función BSU1, BIN2 siempre estará en el estado ON e inhibirá las actividades de BZR1/2, disminuyendo así la respuesta a los BR.
Actividad 3
1. Predecir el fenotipo de una ganancia de función BSK.
- crecimiento no regulado
- sin efecto
- enano
- no puede predecir
Respuesta: a. BSK es un regulador positivo de la vía que funciona para activar BSU1. Por lo tanto, una ganancia de función provocaría que activara positivamente la vía, incluso en ausencia de hormona BR, provocando un crecimiento no regulado.
2. Ahora predice el fenotipo de un mutante doble entre la ganancia de función de BSK y una ganancia de función de BIN2.
- crecimiento no regulado
- sin efecto
- enano
- no puede predecir
Respuesta: c. La mutación BIN2 de ganancia de función bloquea BIN2 en el estado ON donde funcionaría inhibiendo la actividad de BZR1/2. Sería impermeable a la regulación por el BSU1 aguas arriba, por lo que incluso si BSU1 estuviera continuamente activo debido a la ganancia de función BSK, el mutante BIN2 inhibiría el crecimiento y por lo tanto sería epistático.
Ahora intentas un par. Rellena cada una de las siguientes tablas para determinar si la Actividad hipotética será activada o reprimida en respuesta a cada Estímulo.
Referencias
Sharma, M., Cortes-Cruz, M., Ahern, K.R., McMullen, M., Bruknell, T.P., y Chopra, S. (2011). La identificación del producto del gen Pr1 completa la vía de biosíntesis de antocianinas del maíz. Genética 188, 69-79.