
1.12: Ingeniería Genética
- Page ID
- 56629

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)- Definir ingeniería genética.
- Enumere y explique brevemente los cinco pasos básicos de la ingeniería genética. Describir por qué cada uno es necesario.
- Identificar las diferencias fundamentales entre los cultivos genéticamente modificados y los cultivos no modificados genéticamente.
- Explicar las limitaciones a la cría tradicional que son superadas por la ingeniería genética.
- Identificar el tiempo aproximado requerido para obtener una línea de cultivo transgénico comercializable (completar todo el proceso de ingeniería genética del cultivo).
Introducción
La producción de plantas genéticamente modificadas se hizo posible después de que Bob Fraley y otros lograron utilizar Agrobacterium tumefaciens para transformar células vegetales con ADN recombinante a principios de la década de 1980 (Vasil, 2008a). Desde este avance en la biotecnología vegetal, los cultivos transgénicos ahora se desarrollan y cultivan rutinariamente en muchas partes del mundo. Las estadísticas actuales sobre la adopción de cultivos genéticamente modificados en Estados Unidos se pueden encontrar en el sitio web del Servicio de Investigación Económica del USDA.
La ingeniería genética se ha utilizado con éxito para desarrollar nuevos genes de importancia económica que pueden ser utilizados para mejorar la genética de las plantas de cultivo. La ingeniería genética es la adición dirigida de un gen o genes extraños al genoma de un organismo. Los genes pueden aislarse de un organismo y transferirse a otro o pueden ser genes de una especie que se modifican y reinsertan en la misma especie. Los nuevos genes, comúnmente denominados transgenes, se insertan en una planta mediante un proceso llamado transformación. El gen insertado contiene información que le dará al organismo un rasgo (Figura 1).
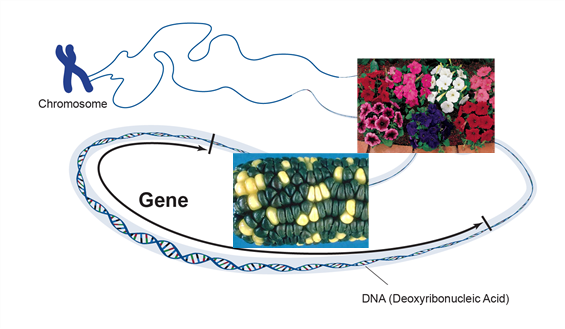
La mejora genética de cultivos (fitomejoramiento) es una herramienta importante pero tiene limitaciones. Primero, en términos convencionales, la mejora genética solo se puede hacer entre dos plantas que pueden aparearse sexualmente entre sí. Esto limita los nuevos rasgos que se pueden sumar a los que ya existen en esa especie. Segundo, cuando las plantas se aparean, (cruzan), muchos rasgos se transfieren junto con el rasgo de interés, incluyendo rasgos con efectos indeseables sobre el potencial de rendimiento.
La ingeniería genética, por otro lado, no está vinculada por estas limitaciones. Elimina físicamente el ADN de un organismo y transfiere el (los) gen (es) para uno o algunos rasgos a otro. Dado que el cruce no es necesario, se supera la barrera 'sexual' entre especies. Por lo tanto, los rasgos de cualquier organismo vivo pueden transferirse a una planta. Este método es más específico ya que se puede agregar un solo rasgo a una planta.
El proceso general de ingeniería genética. Se proporciona una explicación básica de los cinco pasos para la ingeniería genética de un cultivo. Los cinco pasos son:
- Localizar un organismo con un rasgo específico y extraer su ADN.
- Clonar un gen que controle el rasgo.
- Diseñar un gen para expresarse de una manera específica.
- Transformación, insertando el gen en las células de una planta de cultivo.
- Cruzar el transgén en un fondo de élite.
Paso 1: Extracción de ADN
El proceso de ingeniería genética requiere la finalización exitosa de una serie de cinco pasos y descubrimientos. Para entender mejor cada uno de estos, se utilizará como ejemplo el desarrollo del maíz Bt.
Antes de que pueda comenzar el proceso de ingeniería genética, se debe descubrir un organismo vivo que exhiba el rasgo deseado. El rasgo del maíz Bt (resistencia al barrenador europeo del maíz) fue descubierto hace unos 100 años. Los agricultores de gusanos de seda en Oriente habían notado que las poblaciones de gusanos de seda estaban muriendo. Los científicos descubrieron que una bacteria natural del suelo estaba causando la muerte del gusano de seda. Estas bacterias del suelo, llamadas Bacillus thuringiensis, o Bt para abreviar, produjeron una proteína que era tóxica para los gusanos de seda, la proteína Bt.
Aunque los científicos no lo sabían, habían hecho uno de los primeros descubrimientos necesarios en el proceso de elaboración del maíz Bt. La misma proteína Bt que se encuentra tóxica para los gusanos de seda también es tóxica para el barrenador del maíz europeo porque ambos insectos pertenecen al orden de los Lepidópteros. La producción de la proteína Bt en la bacteria está controlada por los genes de la bacteria.
Para poder trabajar con el gen responsable de elaborar la toxina Bt, los científicos deben extraer ADN de la bacteria Bt (Figura 2). Esto se logra tomando una muestra de bacterias que contienen el gen de interés y llevándolo a través de una serie de pasos que separan el ADN de las otras partes de una célula.
Paso 2: Clonación de genes
El segundo paso del proceso de ingeniería genética es la clonación de genes. Durante la extracción de ADN, todo el ADN del organismo se extrae a la vez. Esto significa que la muestra de ADN extraída de la bacteria Bacillus thuringiensis contendrá el gen para la proteína Bt, pero también todos los genes de la otra bacteria. Los científicos utilizan la clonación de genes para separar el gen único de interés del resto del ADN extraído (Figura 2).
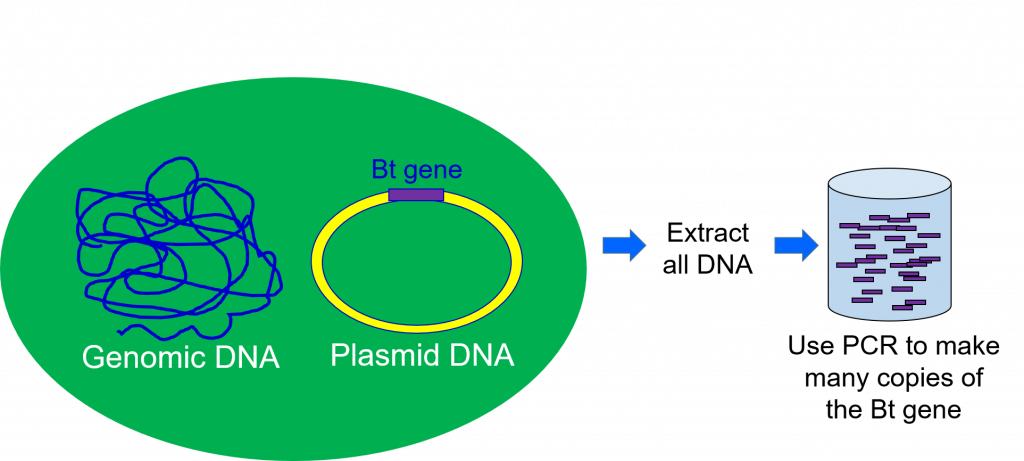
Las siguientes etapas de la ingeniería genética implicarán un mayor estudio y experimentación con este gen. Para ello, un científico necesita tener miles de copias exactas del mismo. Esta copia también se realiza durante la etapa de clonación génica.
Paso 3: Diseño de genes
El diseño de genes se basa en otro descubrimiento importante. Esta fue la teoría de 'Un gen Una enzima' propuesta por primera vez por George W. Beadle y Edward L. Tatum en la década de 1940. Los descubrimientos realizados durante su investigación sentaron las bases para la teoría de que un solo gen almacena la información que dirige a la célula en cómo producir una sola enzima (proteína). Por lo tanto, existe un solo gen que controla la producción de la proteína Bt. Se llama el gen Bt.
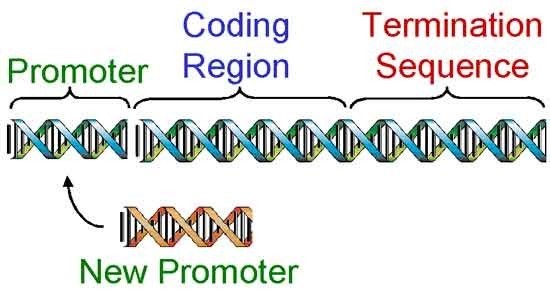
Una vez clonado un gen (Figura 2), los ingenieros genéticos inician el tercer paso, diseñando el gen para que funcione una vez dentro de un organismo diferente. Esto se realiza en un tubo de ensayo cortando el gen con enzimas de restricción y reemplazando ciertas regiones (Figura 3).
Los científicos reemplazaron el promotor del gen bacteriano con promotores que activan el gen Bt en partes seleccionadas de la planta o promotores que siempre pueden activar el gen Bt en todos los tejidos. Como resultado, el primer gen Bt liberado fue diseñado para producir un nivel de proteína Bt letal para el barrenador europeo del maíz y para producir únicamente la proteína Bt en tejidos verdes de la planta de maíz, (tallos, hojas, etc.). Posteriormente, se diseñaron genes Bt para producir el nivel letal de proteína en todos los tejidos de una planta de maíz, (hojas, tallos, borla, oreja, raíces, etc.).
Transformación de plantas y cultivo de tejidos
El proceso de transformación implica la inserción del constructo transgénico deseado (Figura 5) en células de la especie vegetal receptora. En este proceso, los científicos aíslan tejido o células del cultivar que desean transformar y utilizan uno de varios métodos para insertar el transgén en el tejido o células. La construcción transgénica contiene las siguientes características clave.
- Un promotor que actúa para activar y desactivar el gen en la célula. El promotor CaMV 35s del virus del mosaico de la coliflor (CaMV) se usa comúnmente en ingeniería genética. Otros tipos de promotores, como el promotor de la nopalina sintasa (NOS-pro) también se pueden usar para expresar transgenes en tejidos vegetales.
- Un marcador seleccionable que se utiliza para seleccionar células que obtuvieron exitosamente el constructo durante el proceso de transformación. En la figura 4, el marcador seleccionable en el constructo es NPT II (Kanr) que controla la resistencia al antibiótico kanamicina. Las células de la planta utilizadas para la transformación se cultivarán en un medio que contenga el antibiótico. Otros marcadores seleccionables que se han utilizado con éxito en plantas incluyen genes que controlan la resistencia a herbicidas.
- Se incluye una secuencia terminadora, como la nopalina sintasa (NOS) para marcar el final de la secuencia transgénica para una expresión adecuada en células vegetales.

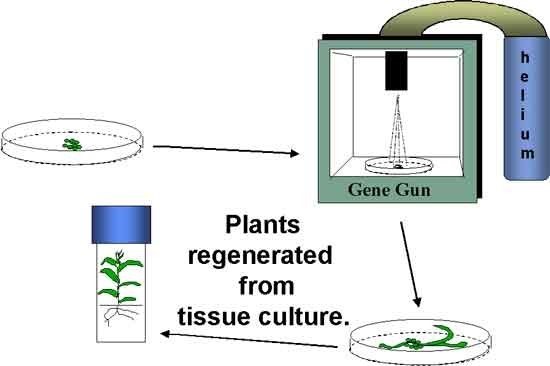
Dos métodos de transformación comúnmente utilizados incluyen la transformación mediada por Agrobacterium tumefaciens y la transformación biolística (también conocida como pistola génica), comúnmente conocida como bombardeo de partículas (Figura 5). El método biolístico implica el uso de alta presión para impulsar perlas de tungsteno u oro recubiertas con ADN del constructo génico hacia las células vegetales.
Transformación de plantas mediada por Agrobacterium
Las agallas coronarias son tumores de plantas que surgen en el sitio de infección por algunas especies de Agrobacterium. Las agrobacterias no ingresan a las células vegetales sino que transfieren un segmento de ADN llamado ADN-T desde su plásmido circular extracromosómico inductor de tumores (Ti) al genoma de las células hospedadoras. Los plásmidos Ti se mantienen en Agrobacterias debido a que una parte de su ADN-T contiene genes que codifican aminoácidos inusuales utilizados por Agrobacterium. El ADN-T también codifica genes que afectan a la fisiología hormonal de la planta hospedadora dando como resultado el crecimiento inducido de las células infectadas y la formación de tumores. Los científicos aprovecharon la capacidad de Agrobacterium para integrar de manera estable su ADN-T en el genoma de la planta para introducir ADNr en las células vegetales. Primero eliminaron los genes que causan el tumor o la enfermedad de la agalla de la corona en las plantas del ADN-T y diseñaron el plásmido para su replicación tanto en células de Escherichia coli como en Agrobacterium. La replicación inicial del constructo en E. coli es útil para verificar la presencia del gen clonado y aumentar la cantidad de ADN del constructo para usos posteriores, incluyendo secuenciación y transformación en Agrobacterium.
Los pasos en la transformación mediada por Agrobacterium de plantas se describen en la Figura 6.
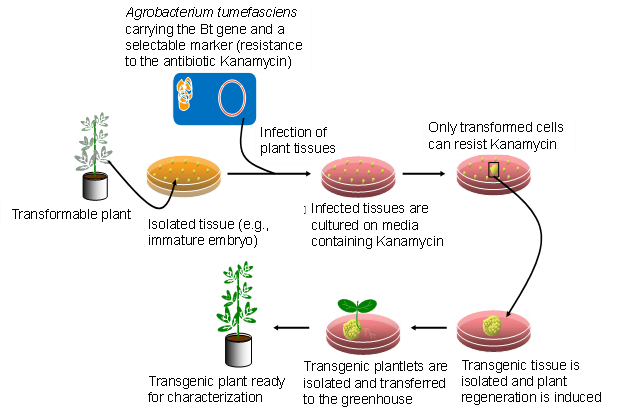
En la actualidad, muy pocas células hospedadoras reciben el constructo durante el proceso de transformación. Cada inserción aleatoria del constructo en el genoma de las células vegetales se denomina evento. Los eventos útiles son raros debido a la naturaleza aleatoria del proceso de transformación. Los marcadores seleccionables son muy importantes porque permiten la identificación de los eventos raros (Figura 7). Los científicos deben seleccionar muchos transformantes potenciales para identificar eventos que sean útiles para la cría.
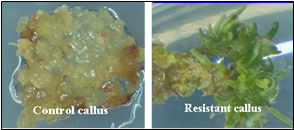
A partir de ahí, el nuevo ADN puede o no insertarse con éxito en un cromosoma. Las células que sí reciben el nuevo gen se denominan transgénicas y se seleccionan de aquellas que no son transgénicas (Figura 7). Muchos tipos de células vegetales son totipotentes, lo que significa que una sola célula vegetal puede convertirse en una planta completa. Por lo tanto, cada célula transgénica puede entonces desarrollarse en una planta completa que tiene el transgén en cada célula. Las plantas transgénicas se cultivan hasta la madurez en invernaderos y se recolecta la semilla que producen, la cual ha heredado el transgén. El trabajo del ingeniero genético ya está completo. Entregará las semillas transgénicas a un obtentor que es responsable del paso final.
Herencia de un transgén en plantas
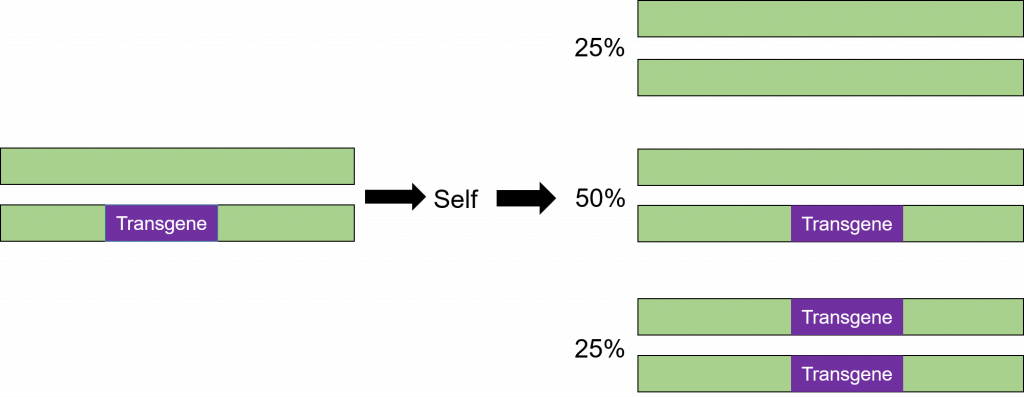
La transformación es exitosa cuando se incorpora un transgén a uno de los cromosomas. Se dice que las células que tienen una sola copia del transgén en sus genomas son hemicigóticas (hemi = mitad, cigoto = cigoto). Debido a que la segregación en la progenie de una planta hemicigótica es la misma que para una planta heterocigótica, el término heterocigótica se utilizará en este curso al referirse a una planta que no es homocigótica para el transgén. El rasgo se segregará en la progenie de la misma manera que cualquier otro gen en la planta como se ilustra a continuación (Figura 8).
Paso 5: Reproducción de retrocruzamiento
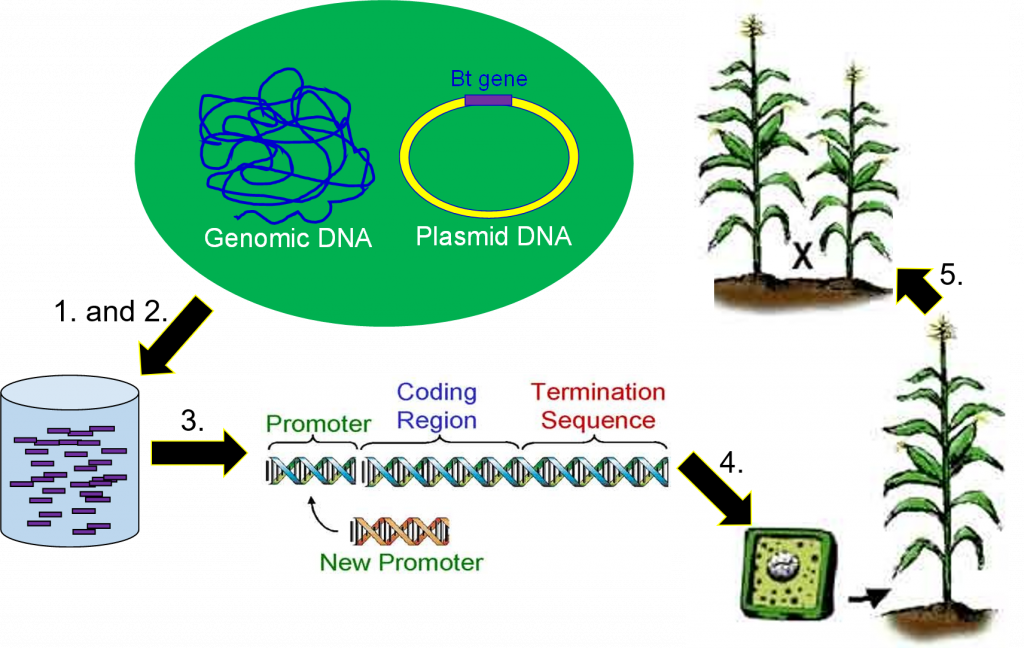
La quinta y última parte de la producción de un cultivo genéticamente modificado es la reproducción por retrocruzamiento (Figura 9). Las plantas transgénicas se cruzan con líneas de reproducción de élite utilizando métodos tradicionales de mejoramiento de plantas para combinar los rasgos deseados de los padres élite y el transgén en una sola línea. Las crías se cruzan repetidamente de regreso a la línea élite para obtener una línea transgénica de alto rendimiento. El resultado será una planta con un potencial de rendimiento cercano a los híbridos actuales que expresa el rasgo codificado por el nuevo transgén.

El Proceso de Ingeniería Genética Vegetal
Todo el proceso de ingeniería genética es básicamente el mismo para cualquier planta. El tiempo requerido para completar los cinco pasos de principio a fin varía según el gen, las especies de cultivo y los recursos disponibles. Puede tomar entre 6 y 15 años más antes de que un nuevo híbrido transgénico esté listo para su liberación para ser cultivado en campos de producción.
El proceso de cultivo tisular de regeneración de plantas transgénicas a partir del callo puede resultar en una variación genética que no está asociada con el transgén. Además, la línea padre utilizada para la transformación comúnmente se selecciona por la frecuencia con la que se pueden obtener eventos útiles y no por su desempeño agronómico. Por lo tanto, los transgenes se incorporan a cultivares comerciales mediante procedimientos de mejoramiento convencionales, como el retrocruzamiento.
La ingeniería genética es la adición dirigida de ADN extraño (genes) en un organismo.
Cinco pasos básicos en la ingeniería genética de cultivos:
- Extracción de ADN — El ADN se extrae de un organismo que se sabe que tiene el rasgo deseado.
- Clonación de genes — El gen de interés es localizado y copiado.
- Modificación génica — El gen se modifica para expresarse de una manera deseada alterando y reemplazando las regiones génicas.
- Transformación — El gen o genes se entregan en células de cultivo tisular, utilizando uno de varios métodos, donde ojalá aterricen en el núcleo y se inserten en un cromosoma.
- Reproducción de retrocruzamiento — Las líneas transgénicas se cruzan con líneas de élite para hacer líneas transgénicas de alto
Referencias
Vasil, I. K. (2008) Una breve historia de la biotecnología vegetal. Phytochem 7:387-394.
Vasil, I. K. (2008) Una historia de la biotecnología vegetal: de la Teoría Celular de Shleiden y Schwann a los cultivos biotecnológicos. Célula Vegetal Rep 27:1423-1440.