
2.2: Alelos múltiples, dominancia incompleta y codominancia




- Última actualización
- 29 oct 2022
- Guardar como PDF
( \newcommand{\kernel}{\mathrm{null}\,}\)
Introducción
Gregor Mendel supo mantener las cosas simples. En el trabajo de Mendel sobre plantas de guisante, cada gen vino en solo dos versiones diferentes, o alelos, y estos alelos tenían una relación de dominación agradable y clara (con el alelo dominante anulando completamente al alelo recesivo para determinar la apariencia de la planta).
Hoy en día, sabemos que no todos los alelos se comportan tan directamente como en los experimentos de Mendel. Por ejemplo, en la vida real:
-
Los pares de alelos pueden tener una variedad de relaciones de dominio (es decir, un alelo del par puede no “ocultar” completamente al otro en el heterocigoto).
-
A menudo hay muchos alelos diferentes de un gen en una población.
En estos casos, el genotipo de un organismo, o conjunto de alelos, aún determina su fenotipo, o características observables. Sin embargo, una variedad de alelos pueden interactuar entre sí de diferentes maneras para especificar el fenotipo.
Como nota al margen, probablemente tengamos suerte de que los genes de guisante de Mendel no mostraran estas complejidades. Si lo hubieran hecho, es posible que Mendel no hubiera entendido sus resultados, y no hubiera descubierto los principios básicos de la herencia, ¡que son clave para ayudarnos a entender los casos especiales!
Dominio incompleto
Los resultados de Mendel fueron pioneros en parte porque contradijeron la idea (entonces popular) de que los rasgos de los padres se mezclaban permanentemente en sus crías. En algunos casos, sin embargo, el fenotipo de un organismo heterocigótico en realidad puede ser una mezcla entre los fenotipos de sus progenitores homocigóticos.
Por ejemplo, en la boca de dragón, Antirrhinum majus, un cruce entre una planta homocigótica de flores blancas (CWCW) y una planta homocigótica de flores rojas (CRCR) producirá descendencia con flores rosadas (CRCW). Este tipo de relación entre alelos, con un fenotipo heterocigoto intermedio entre los dos fenotipos homocigotos, se denomina dominancia incompleta.
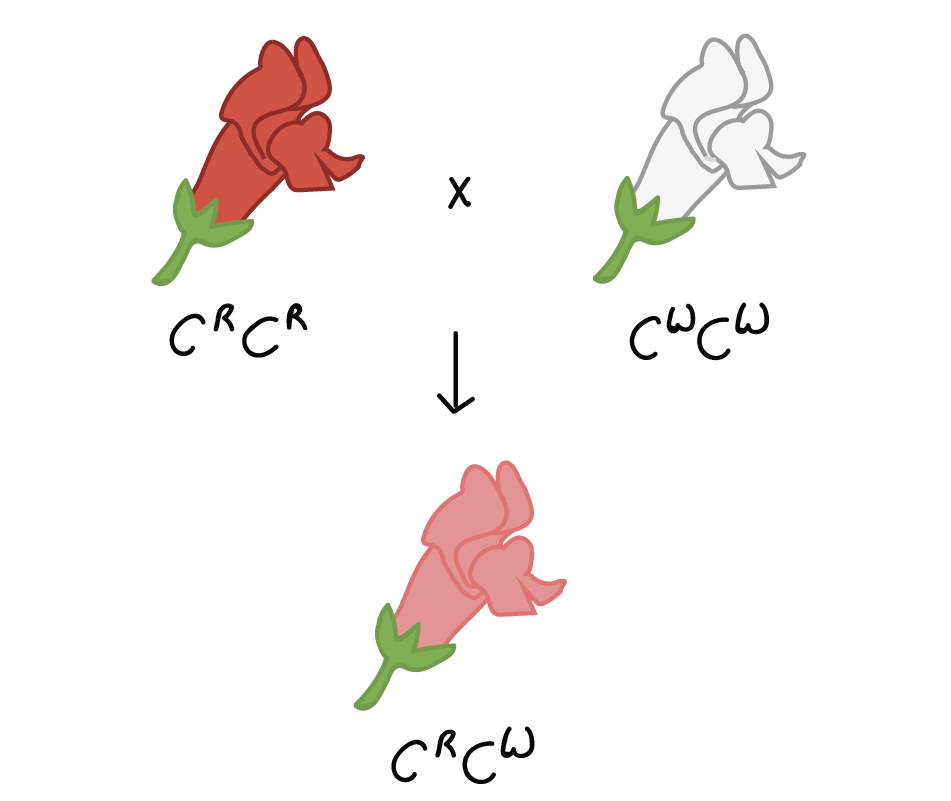
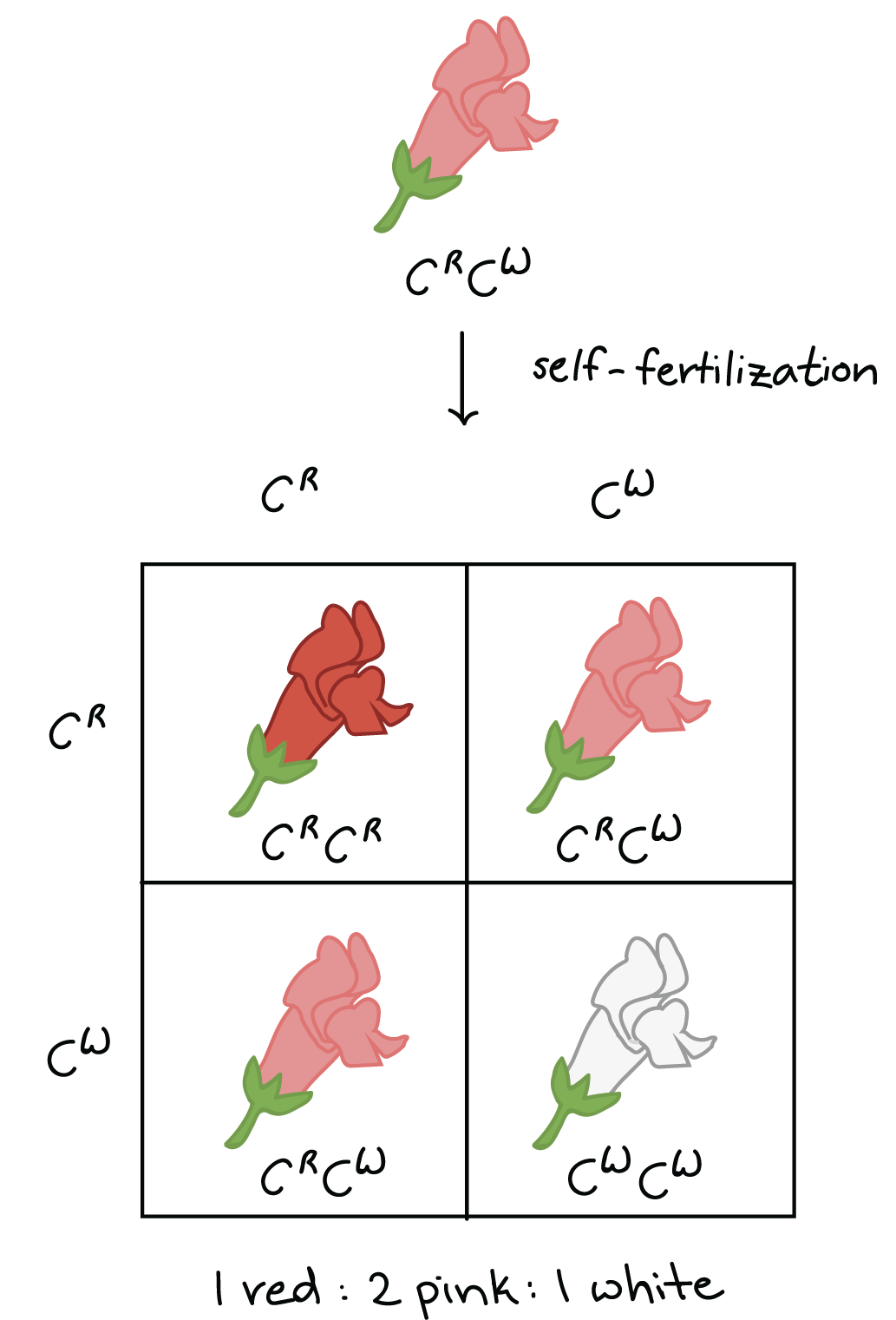
Todavía podemos usar el modelo de Mendel para predecir los resultados de cruces para alelos que muestran dominancia incompleta. Por ejemplo, la autofertilización de una planta rosada produciría una relación genotipo1CRCR:2CRCW:1CWCW y una relación de fenotipo de1:2:1 rojo:rosa:blanco. Los alelos aún se heredan de acuerdo con las reglas básicas de Mendel, incluso cuando muestran un dominio incompleto.
Codominancia
Estrechamente relacionado con la dominancia incompleta está la codominancia, en la que ambos alelos se expresan simultáneamente en el heterocigoto.
Podemos ver un ejemplo de codominancia en los grupos sanguíneos MN de los humanos (menos famosos que los grupos sanguíneos ABO, ¡pero aún importantes!). El tipo de sangre MN de una persona está determinado por sus alelos de cierto gen. UnLM alelo especifica la producción de un marcador M que se muestra en la superficie de los glóbulos rojos, mientras que unLN alelo especifica la producción de un marcador N ligeramente diferente.
Los homocigotos (LMLMyLNLN) solo tienen marcadores M o N, respectivamente, en la superficie de sus glóbulos rojos. Sin embargo, los heterocigotos (LMLN) tienen ambos tipos de marcadores en números iguales en la superficie celular.
En cuanto al dominio incompleto, todavía podemos usar las reglas de Mendel para predecir la herencia de alelos codominantes. Por ejemplo, si dos personas conLMLN genotipos tuvieran hijos, esperaríamos ver los tipos de sangre M, MN y N yLMLM,LMLN, yLNLN genotipos en sus hijos en una1:2:1 proporción (¡si tuvieran suficientes hijos para que determinemos las proporciones con precisión!)
Alelos múltiples
El trabajo de Mendel sugirió que solo existían dos alelos por cada gen. Hoy, sabemos que ese no es siempre, ni siquiera usualmente, ¡el caso! Aunque los humanos individuales (y todos los organismos diploides) solo pueden tener dos alelos para un gen dado, pueden existir múltiples alelos en un nivel poblacional, y diferentes individuos en la población pueden tener diferentes pares de estos alelos.
Como ejemplo, consideremos un gen que especifica el color del pelaje en los conejos, llamadoC gen. ElC gen viene en cuatro alelos comunes:C,cch,ch, yc:
- UnCC conejo tiene pelaje negro o marrón.
- Uncchcch conejo tiene coloración chinchilla (pelaje grisáceo).
- Unchch conejo tiene patrones del Himalaya (punto de color), con un cuerpo blanco y orejas oscuras, cara, pies y cola.
- Uncc conejo es albino, con una bata blanca pura.
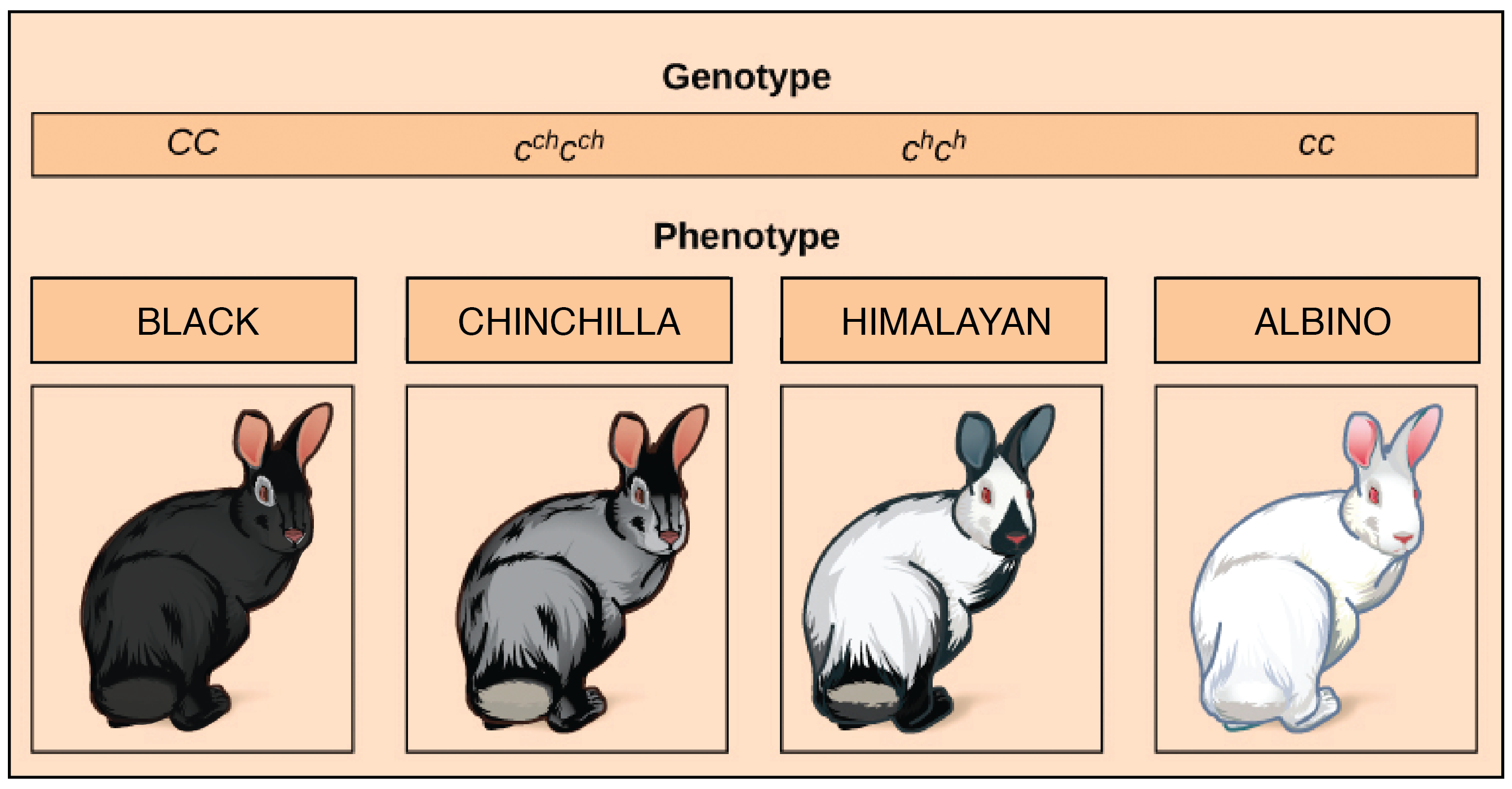
Los alelos múltiples hacen que muchas relaciones de dominación sean posibles. En este caso, elC alelo negro es completamente dominante a todos los demás; elcch alelo chinchilla es incompletamente dominante a losc alelos del Himalayach y albina; y elch alelo del Himalaya es completamente dominante alc alelo alino.
Los criadores de conejos descubrieron estas relaciones cruzando diferentes conejos de diferentes genotipos y observando los fenotipos de los kits heterocigotos (conejitos bebés).
- [¿Cómo cambian estos alelos el color del conejo?]
-
ElC gen en conejos codifica una enzima que es necesaria para elaborar un tipo de pigmento llamado melanina en los pelos 1,2.
- ElC alelo de este gen codifica una enzima completamente funcional que produce mucho pigmento y da como resultado pelaje negro.
- Elcch alelo codifica una enzima que es menos efectiva en la fabricación de pigmento, dando como resultado un pelaje más claro y más grisáceo.
- Elch alelo codifica una enzima defectuosa, donde el defecto hace que la enzima sea muy sensible a la temperatura: funciona bien a bajas temperaturas, pero no funciona en absoluto a temperaturas más altas. Las extremidades del conejo (patas, orejas, etc.) son más frías, por lo que la enzima funciona ahí y hace pigmento. El cuerpo principal del conejo es más cálido, por lo que la enzima no funciona y no se elabora ningún pigmento.
- Elc alelo codifica una enzima completamente no funcional, lo que lleva a un conejo albino (uno que no produce ningún pigmento en sus pelos).
Nota para los colombóreos del conejo: El color del pelaje del conejo está determinado por una serie de genes, no solo por elC gen. La serie alélica aquí descritaC asume un cierto trasfondo genético para los otros genes, uno en el que unCC genotipo da como resultado un conejo negro.
Colaboradores y Atribuciones
- [Atribución y referencias]
-
Atribución:
Este artículo es un derivado modificado de “Características y rasgos”,” por OpenStax College, Biology (CC BY 3.0). Descarga gratis el artículo original en http://cnx.org/contents/185cbf87-c72e-48f5-b51e-f14f21b5eabd@9.85.
El artículo modificado está licenciado bajo una licencia CC BY-NC-SA 4.0.
Obras citadas:
- Color de pelaje en el conejo del Himalaya. (n.d.). En Cengage learning. Recuperado de http://www.cengage.com/biology/discipline_content/animations/himalayan_rabbit_m.html.
- Hinkle, Amy. (n.d.). Bioquímica del color de la capa de conejo. En el rancho de conejos de Amy. Recuperado de http://www.amysrabbitranch.com/Color%26Genetics/Biochemistry-CoatColor.pdf.
Referencias adicionales:
Blumberg, R. B. (1997). Ponencia de Mendel en inglés. En MendelWeb. Recuperado de http://www.mendelweb.org/Mendel.plain.html.
Coat color photo matrix. (2013, 29 de septiembre) .In Granja granero verde. Recuperado de http://www.gbfarm.org/rabbit/holland-colors-matrix.shtml.
Genética del color: La serie C. (n.d.) En El sendero natural. Recuperado de http://www.thenaturetrail.com/rabbit-genetics/color-c-series-chinchilla-sable-himalayan-rew/.
Regulador de conductancia transmembrana de fibrosis quística. (2015, 12 de noviembre). Recuperado el 22 de noviembre del 2015 de Wikipedia: https://en.wikipedia.org/wiki/Cystic_fibrosis_transmembrane_conductance_regulator.
Dominancia (genética). (2015, 3 de noviembre). Recuperado el 21 de noviembre del 2015 de Wikipedia: https://en.wikipedia.org/wiki/Dominance_%28genetics%29.
Fox, Richard R. (1974). Taxonomía y genética. En S. H. Weisbroth, R. E. Flatt, y A. L. Kraus (Eds.), La biología del conejo de laboratorio (6-22). Nueva York, NY: Prensa Académica.
Orfano, Finn. (2010, 15 de diciembre). La codominancia en genética: Una visión general. En buje Bright. Recuperado de http://www.brighthub.com/science/genetics/articles/99400.aspx.
Purves, W. K., Sadava, D., Orians, G. H., y Heller, H. C. (2003). Alelos y sus interacciones. En la vida: La ciencia de la biología (7ª ed., pp. 197-199). Sunderland, MA: Asociados Sinauer.
Tabla de genotipos de color de conejo. (n.d.). En El sendero natural. http://www.thenaturetrail.com/rabbit-genetics/rabbit-color-genotypes-chart/.
Raven, P. H., Johnson, G. B., Mason, K. A., Losos, J. B., y Singer, S. R. (2014). Patrones de herencia. En Biología (10a ed., AP ed., pp. 221-238). Nueva York, NY: McGraw-Hill.
Reece, J. B., Urry, L. A., Caín, M. L., Wasserman, S. A., Minorsky, P. V., y Jackson, R. B. (2011). Mendel y la idea genética. En Campbell Biology (10a ed., pp. 267-291). San Francisco, CA: Pearson.
Prueba de sudor. (2015, Septebmer 24). Recuperado el 22 de noviembre del 2015 de Wikipedia: https://en.wikipedia.org/wiki/Sweat_test.