
4: Energía y Ecosistemas



- Última actualización
- 30 oct 2022
- Guardar como PDF

( \newcommand{\kernel}{\mathrm{null}\,}\)
Objetivos de aprendizaje
Después de completar este capítulo, podrás
- Describir la naturaleza de la energía, sus diversas formas y las leyes que rigen sus transformaciones.
- Explicar cómo la Tierra es un sistema de flujo continuo para la energía solar.
- Identificar los tres componentes principales del presupuesto energético de la Tierra.
- Describir las relaciones energéticas dentro de los ecosistemas, incluyendo la fijación de la energía solar por los productores primarios y el paso de esa energía fija a través de otros componentes del ecosistema.
- Explicar por qué la estructura trófica de la productividad ecológica tiene forma de pirámide y por qué los ecosistemas no pueden soportar a muchos depredadores superiores.
- Compara las estrategias de alimentación de los humanos que viven un estilo de vida de caza y recolección con las de la gente urbana moderna.
Introducción
Ninguno del planeta Tierra, su biosfera, o ecosistemas a ninguna escala son autosustentables con respecto a la energía. De hecho, sin un acceso continuo a una fuente externa de energía, todas estas entidades agotarían rápidamente sus cantidades de energía almacenada y se enfriarían rápidamente, y en el caso de la biosfera y los ecosistemas, dejarían de funcionar de formas que sustentan la vida. La fuente externa de energía a esos sistemas es la energía solar, que se almacena principalmente como calor y biomasa. En efecto, la energía solar es absorbida por las plantas verdes y las algas y se utiliza para fijar el dióxido de carbono y el agua en azúcares simples a través de un proceso conocido como fotosíntesis. Esta fijación biológica de la energía solar proporciona la base energética para casi todos los organismos y ecosistemas (las pocas excepciones se describen más adelante). La energía es crítica para el funcionamiento de los procesos físicos en todo el universo, y de los procesos ecológicos en la biosfera de la Tierra. En este capítulo examinaremos la naturaleza física de la energía, las leyes que rigen su comportamiento y transformaciones, y su papel en los ecosistemas.
La naturaleza de la energía
La energía es una entidad física fundamental y se define simplemente como la capacidad de un cuerpo o sistema para realizar el trabajo. En física, el trabajo se define como el resultado de una fuerza que se aplica sobre una distancia. En todos los siguientes ejemplos de trabajo, la energía se transforma y se logra algún resultado medible:
- Un palo de hockey golpea un disco, lo que hace que se acelere hacia un objetivo
- Se recoge un libro del piso, se levanta y luego se coloca sobre una mesa
- Un vehículo es conducido a lo largo de una carretera
- El calor de una estufa es absorbido por el agua en una tetera, lo que hace que se caliente y eventualmente hierva
- El pigmento fotosintético clorofila absorbe la luz solar, convirtiendo la energía electromagnética en una forma que las plantas y las algas pueden utilizar para sintetizar azúcares
La energía puede existir en diversos estados, cada uno de los cuales es fundamentalmente diferente de los demás. Sin embargo, bajo condiciones adecuadas la energía en cualquier estado puede convertirse en otro a través de transformaciones físicas o químicas. Los estados de energía pueden agruparse en tres categorías: electromagnética, cinética y potencial.
Energía Electromagnética
La energía electromagnética (o radiación electromagnética) está asociada con fotones. Estas tienen propiedades tanto de partículas como de ondas y viajan por el espacio a una velocidad constante de 3 × 10 8 m/s (la velocidad de la luz). La energía electromagnética existe como un espectro continuo de longitudes de onda, que (ordenadas de las longitudes de onda más cortas a más largas) se conocen como gamma, rayos X, ultravioleta, luz visible, infrarroja, microondas y radio (Figura 4.1). El ojo humano puede percibir energía electromagnética en un rango de longitudes de onda de aproximadamente 0.4 a 0.7 μm, una parte del espectro que se denomina radiación visible o luz (1 μm, o 1 micrómetro, es de 10 —6 m; ver Apéndice A).
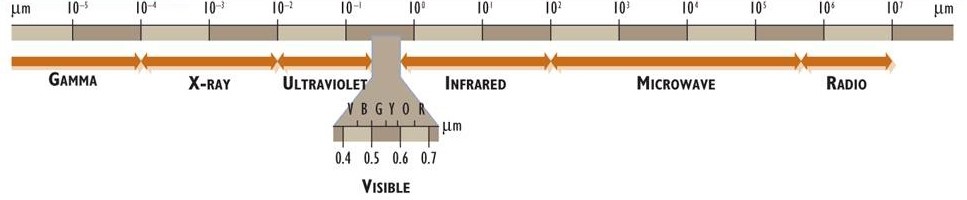
La energía electromagnética es emitida (o radiada) por todos los objetos que tienen una temperatura superficial superior a cero absoluto (mayor a -273°C o 0°K). La temperatura superficial de un cuerpo determina la velocidad y la calidad espectral de la radiación que emite. En comparación con un cuerpo más frío, uno más caliente tiene una tasa de emisión mucho mayor, y la radiación está dominada por longitudes de onda más cortas y de mayor energía. Por ejemplo, el Sol tiene una temperatura superficial extremadamente caliente de aproximadamente 6000°C, y como consecuencia directa la mayor parte de la radiación que emite es ultravioleta (0.2 a 0.4 μm), visible (0.4 a 0.7 μm) y cerca del infrarrojo (0.7 a 2 μm). (Tenga en cuenta que el interior del Sol es mucho más caliente que 6000°C, pero es la temperatura de la superficie la que influye directamente en la radiación emitida). Debido a que la temperatura superficial de la Tierra promedia mucho más fría a aproximadamente 15°C, irradia cantidades mucho más pequeñas de energía a longitudes de onda más largas (alcanzando un pico a una longitud de onda de aproximadamente 10 μm).

Energía cinética
La energía cinética se asocia con cuerpos que están en movimiento. Se pueden distinguir dos clases de energía cinética.
La energía cinética mecánica se asocia con cualquier objeto que esté en movimiento, lo que significa que viaja de un lugar a otro. Por ejemplo, un disco de hockey que vuela por el aire, una bicicleta de sendero que se monta por un sendero, un ciervo que corre por un bosque, el agua que fluye en un arroyo o un planeta que se mueve por el espacio son expresiones de este tipo de energía cinética. La cantidad de energía cinética mecánica está determinada por la masa de un objeto y su velocidad.
La energía cinética térmica se asocia con la velocidad a la que los átomos o moléculas están vibrando. Dichas vibraciones se congelan a -273°C (cero absoluto), pero son progresivamente más vigorosas a temperaturas más altas, lo que corresponde a un mayor contenido de energía cinética térmica, a la que también se le conoce como calor.
Energía Potencial
La energía potencial es la capacidad almacenada para realizar el trabajo. Para realizar realmente el trabajo, la energía potencial debe transformarse en energía electromagnética o cinética. Hay una serie de tipos de energía potencial:
La energía potencial gravitacional es el resultado de la gravedad, o de las fuerzas atractivas que existen entre todos los objetos. Por ejemplo, el agua almacenada a cualquier altura sobre el nivel del mar contiene energía potencial gravitacional. Esto se puede convertir en energía cinética si existe una vía que permita que el agua fluya cuesta abajo. La energía potencial gravitacional se puede convertir en energía eléctrica a través de la tecnología de centrales hidroeléctricas.
La energía potencial química se almacena en los enlaces entre los átomos dentro de las moléculas. La energía potencial química puede ser liberada por reacciones exotérmicas (aquellas que conducen a una liberación neta de energía térmica), como en los siguientes ejemplos:
- La energía potencial química se almacena en los enlaces moleculares de los minerales de sulfuro, como el sulfuro de hierro (FeS 2), y parte de esta energía se libera cuando los sulfuros se oxidan. Las bacterias especializadas pueden aprovechar metabólicamente la energía potencial de los sulfuros para apoyar su propia productividad, a través de un proceso conocido como quimiosíntesis (esto se examina más adelante en este capítulo).
- Los enlaces iónicos de las sales también almacenan energía potencial química. Por ejemplo, cuando el cloruro de sodio (sal de mesa, NaCl) se disuelve en agua, la energía potencial iónica se libera como calor, lo que aumenta ligeramente la temperatura del agua.
- Los hidrocarburos almacenan energía en los enlaces entre sus átomos de hidrógeno y carbono (los hidrocarburos contienen solo estos átomos). La energía potencial química de la gasolina, una mezcla de hidrocarburos líquidos, se libera en un motor de combustión interna y se transforma mecánicamente para lograr la energía cinética del movimiento vehicular.
- Los compuestos orgánicos (bioquímicos) producidos metabólicamente por los organismos también almacenan grandes cantidades de energía potencial en sus enlaces interatómicos. La densidad energética típica de los carbohidratos es de aproximadamente 16.8 kJ/g, mientras que la de las proteínas es 21.0 kJ/g, y los lípidos (o grasas) 38.5 kJ/g Muchos organismos almacenan sus reservas de energía como grasa debido a que estos bioquímicos tienen una densidad energética tan alta.
La energía potencial eléctrica es el resultado de diferencias en la cantidad de electrones, que son partículas subatómicas cargadas negativamente que fluyen de áreas de alta densidad a áreas donde es menor. Cuando se usa un interruptor eléctrico para completar un circuito que conecta dos áreas con diferentes potenciales eléctricos, los electrones fluyen a lo largo del gradiente de electrones. La energía eléctrica puede entonces transformarse en usos como luz, calor o trabajo realizado por una máquina. Una diferencia en el potencial eléctrico se conoce como voltaje, y la corriente de los electrones debe fluir a través de un material conductor, como un metal.
La elasticidad es una especie de energía potencial que es inherente a las cualidades físicas de ciertos materiales flexibles y que puede realizar trabajo cuando se libera, como ocurre cuando se usa un arco dibujado para disparar una flecha.
Los gases comprimidos también almacenan energía potencial, que puede funcionar si se permite que se produzca la expansión. Este tipo de energía potencial está presente en un cilindro que contiene gas comprimido o licuado.
La energía potencial nuclear es el resultado de las fuerzas de unión extremadamente fuertes que existen dentro de los átomos. Esta es, con mucho, la forma de energía más densa. Enormes cantidades de energía electromagnética y cinética se liberan cuando las reacciones nucleares convierten la materia en energía. Una reacción de fisión implicó la división de isótopos de ciertos átomos pesados, como el uranio-235 y el plutonio-239 (U 235 y 239 P), para generar átomos más pequeños más enormes cantidades de energía. Las reacciones de fisión ocurren en explosiones nucleares y, en condiciones controladas, en reactores nucleares utilizados para generar electricidad. Una reacción de fusión implica la combinación de ciertos elementos ligeros, como el hidrógeno, para formar átomos más pesados en condiciones de temperatura y presión extremadamente altas, liberando al mismo tiempo enormes cantidades de energía. Las reacciones de fusión que involucran hidrógeno ocurren en las estrellas y son responsables de las cantidades inimaginablemente grandes de energía que estos cuerpos celestes generan e irradian al espacio. Se piensa que todos los átomos pesados del universo fueron producidos por reacciones de fusión que ocurren en las estrellas (ver Capítulo 3). Las reacciones de fusión también ocurren en un tipo de artefacto explosivo nuclear conocido como bomba de hidrógeno. Aún no se ha desarrollado una tecnología para explotar las reacciones de fusión controlada para generar electricidad; si está disponible, la fusión controlada podría utilizarse para generar cantidades prácticamente ilimitadas de energía comercial (ver Capítulo 13).

Unidades de Energía
Si bien la energía puede existir en diversas formas, todas ellas se pueden medir en unidades iguales o equivalentes. El sistema internacionalmente aceptado para unidades científicas es el sistema SI (Système International d'Unités), y su unidad recomendada para la energía es el joule (J). Un julio se define como la energía requerida para acelerar 1 kg de masa a 1 m/s 2 (1 metro por segundo por segundo) en una distancia de 1 m.
Una caloría (o gramo-caloría, abreviatura cal) es otra unidad de energía. Una caloría equivale a 4.184 J, y se define como la cantidad de energía requerida para elevar la temperatura de 1 g de agua pura en 1°C (específicamente, de 15°C a 16°C). Tenga en cuenta, sin embargo, que la “Calorías” del dietista equivale a 1000 calorías (1 Caloría = 1 kcal). Sin embargo, el contenido energético de muchos productos alimenticios se cotiza ahora en kJ en países que utilizan el sistema SI de unidades, como Canadá.
Transformaciones Energéticas
Como se señaló anteriormente, la energía puede transformarse entre sus diversos estados. Por ejemplo, cuando la radiación electromagnética solar es absorbida por un objeto oscuro, se transforma en energía térmica y el cuerpo absorbente aumenta de temperatura. La energía potencial gravitacional del agua almacenada a una altura se convierte en la energía cinética del agua que fluye en una cascada, que puede aprovecharse utilizando tecnología hidroeléctrica para hacer girar una turbina y generar energía eléctrica. Además, las longitudes de onda visibles de la radiación solar son absorbidas por la clorofila, un pigmento verde en el follaje de las plantas, y parte de la energía capturada se convierte en energía potencial química de los azúcares a través de la bioquímica de la fotosíntesis.
Todas las transformaciones de la energía deben comportarse de acuerdo con ciertos principios físicos, que se conocen como las leyes de la termodinámica. Estos son principios universales, es decir, siempre son ciertos, independientemente de las circunstancias.
La Primera Ley de la Termodinámica
La primera ley de la termodinámica, también conocida como la ley de conservación de la energía, puede afirmarse de la siguiente manera: La energía puede sufrir transformaciones entre sus diversos estados pero nunca se crea ni se destruye; por lo tanto, el contenido energético del universo permanece constante. Una consecuencia de esta ley es que siempre hay un balance cero entre las entradas de energía a un sistema, cualquier almacenamiento neto dentro de él, y la salida de energía del sistema.
Considera el caso de un automóvil circulando por una autopista. El vehículo consume gasolina, un aporte de energía que se puede medir. La energía potencial del combustible se convierte en varios otros tipos de energía, incluida la energía cinética incorporada en el movimiento hacia adelante del vehículo, la energía eléctrica que alimenta las luces y los limpiaparabrisas, el calor de la fricción entre el vehículo y la atmósfera y la superficie de la carretera, y gases de escape calientes ( energía térmica) y combustible no quemado (energía potencial química) que se ventilan a través del tubo de escape. En general, de acuerdo con la primera ley de la termodinámica, una medición precisa de todas estas transformaciones encontraría que, mientras la energía de la gasolina se convirtiera en otras formas diversas, se conservaba la cantidad total de energía (se mantenía constante).
La Segunda Ley de la Termodinámica
La segunda ley de la termodinámica se puede expresar de la siguiente manera: Las transformaciones de la energía pueden ocurrir espontáneamente solo bajo condiciones en las que hay un aumento en la entropía del universo. La entropía es un atributo físico relacionado con el trastorno, y se asocia con el grado de aleatoriedad en las distribuciones de materia y energía. A medida que aumenta la aleatoriedad (trastorno), también lo hace la entropía. Una disminución en el trastorno se conoce como entropía negativa. Considera, por ejemplo, un globo inflado. Debido a la energía potencial de sus gases comprimidos, ese globo puede filtrar lentamente su contenido a la atmósfera circundante, e incluso puede estallar. Cualquiera de estos resultados puede ocurrir de manera espontánea, ya que ambos procesos representarían un incremento en la entropía del universo. Esto se debe a que los gases comprimidos están más ordenados que los ampliamente dispersos en la atmósfera. En contraste, los gases dispersos en la atmósfera nunca se reubicarían espontáneamente para inflar un globo. Un globo solo se inflará si la energía se gasta a través de una aplicación local de trabajo, como por ejemplo por una persona que sopla dentro del globo. Es decir, se debe gastar energía para provocar una disminución local de la entropía en un sistema. No obstante, tenga en cuenta que este costo energético en sí mismo da lugar a un incremento en la entropía del universo. Por ejemplo, el esfuerzo de una persona inflando un globo implica respiración adicional, que utiliza energía bioquímica y da como resultado que se libere calor al ambiente.
Otro ejemplo se refiere al planeta Tierra. El planeta recibe continuamente radiación solar, casi toda la cual es visible y longitudes de onda del infrarrojo cercano en el rango de aproximadamente 0.4 a 2.0 μm. Parte de esta energía electromagnética es absorbida y convertida en energía térmica, la cual calienta la atmósfera y la superficie. El planeta se enfría de la radiación solar absorbida de varias maneras, pero en última instancia esa energía se disipa por una emisión de energía electromagnética al espacio exterior como radiación infrarroja de onda más larga (de una calidad espectral que alcanza su punto máximo a una longitud de onda de 10 μm). En este caso, la radiación solar de longitud de onda relativamente corta se transforma finalmente en la radiación de longitud de onda más larga emitida por la Tierra, un proceso que representa una degradación en la calidad de la energía y un aumento en la entropía del universo.
Un corolario importante (o proposición secundaria) de la segunda ley de la termodinámica es que las transformaciones energéticas nunca pueden ser completamente eficientes; parte del contenido inicial de energía siempre debe convertirse en calor para que la entropía aumente. Esto ayuda a explicar por qué, incluso cuando se utiliza la mejor tecnología disponible, solo alrededor del 30% de la energía potencial de la gasolina se puede convertir en la energía cinética de un automóvil en movimiento, y no más de alrededor del 40% de la energía del carbón o del gas natural puede transformarse en electricidad en una estación generadora. También hay límites termodinámicos para la eficiencia de la fotosíntesis, proceso por el cual las plantas convierten la radiación visible en bioquímica, incluso cuando ocurre en condiciones ideales con suministros óptimos de nutrientes, agua y luz.
Una evaluación superficial podría sugerir que la vida en general parece contradecir la segunda ley de la termodinámica. Las plantas, por ejemplo, absorben longitudes de onda visibles de radiación electromagnética y utilizan esta forma de energía altamente dispersa para fijar moléculas inorgánicas simples (dióxido de carbono y agua) en bioquímicos extremadamente complejos y densos en energía. La biomasa vegetal puede ser consumida por animales y microbios, que sintetizan sus propios bioquímicos complejos. Estas diversas biosíntesis representan transformaciones energéticas que disminuyen en gran medida la entropía local porque la energía electromagnética relativamente dispersa y los compuestos inorgánicos simples se están convirtiendo en los bioquímicos complejos y altamente ordenados de los organismos. ¿Estas transformaciones biológicas contravienen la segunda ley de la termodinámica?
Esta aparente paradoja de la vida puede resolverse utilizando la siguiente lógica: la bioconcentración localizada de la entropía negativa sólo puede ocurrir porque el sistema (refiriéndose en última instancia a la biosfera, o a toda la vida en la Tierra) recibe un aporte constante de energía en forma de radiación solar. Si de alguna manera se terminara esta fuente externa de energía, todos los organismos y materiales orgánicos se degradarían espontáneamente, liberando moléculas inorgánicas simples y calor y aumentando así la entropía del universo. Por lo tanto, la vida y los ecosistemas no pueden sobrevivir sin insumos continuos de energía solar, que se requieren para organizar y mantener su entropía negativa. En este sentido, la biosfera puede verse como una “isla” de entropía negativa, altamente localizada en el espacio y el tiempo, y alimentada continuamente por el Sol como fuente externa de energía.
Tierra: un sistema de flujo de energía
La radiación electromagnética emitida por el Sol es, con mucho, el mayor aporte de energía que impulsa los ecosistemas. La energía solar calienta el planeta, hace circular su atmósfera y sus océanos, evapora su agua y sostiene casi toda su productividad ecológica. Finalmente, toda la energía solar absorbida por la Tierra es re-irradiada de regreso al espacio en forma de radiación electromagnética de una longitud de onda más larga que la capturada originalmente. En otras palabras, la Tierra es un sistema de flujo continuo, con un equilibrio perfecto entre la entrada de energía solar y la salida de energía re-radiada, y sin almacenamiento neto a largo plazo.
Además, casi todos los ecosistemas dependen absolutamente de la radiación solar como fuente de energía que los organismos fotosintéticos (como las plantas y las algas) utilizan para sintetizar compuestos orgánicos simples (como los azúcares) a partir de moléculas inorgánicas (dióxido de carbono y agua). Luego, las plantas y las algas utilizan la energía potencial química en estos azúcares, además de los nutrientes inorgánicos (como el nitrato y el fosfato), para sintetizar una gran diversidad de bioquímicos a través de diversas reacciones metabólicas. Las plantas crecen y se reproducen mediante el uso de estos bioquímicos y su energía potencial. Además, la biomasa vegetal es utilizada como alimento por la enorme cantidad de organismos que son incapaces de fotosíntesis. Estos organismos incluyen herbívoros que comen plantas directamente, carnívoros que comen a otros animales, y detrívoros que se alimentan de biomasa muerta. (Las relaciones energéticas dentro de los ecosistemas se describen más adelante.)
Menos de 0.02% de la energía solar recibida en la superficie de la Tierra es absorbida y fijada por plantas fotosintéticas y algas. Si bien esto representa un componente cuantitativamente trivial del presupuesto energético del planeta, es sumamente importante cualitativamente porque esta energía biológicamente absorbida y fija es la base de la productividad ecológica. Sin embargo, en última instancia y finalmente, la energía solar fijada por las plantas y las algas se libera nuevamente al ambiente como calor y finalmente se irradia de regreso al espacio exterior. Esto refuerza la idea de que la Tierra es un sistema de flujo continuo para la energía, con un baance perfecto entre la entrada y la salida.
Presupuesto Energético de la Tierra
Un presupuesto energético de un sistema describe las tasas de entrada y salida de energía, así como cualquier transformación interna entre sus diversos estados, incluidos los cambios en las cantidades almacenadas. La Figura 4.2 ilustra aspectos clave del presupuesto de energía física de la Tierra.
La tasa de entrada de radiación solar a la Tierra promedia alrededor de 8.36 J/cm 2 -minuto (2.00 cal/cm 2 -min), medida en el límite exterior de la atmósfera. Aproximadamente la mitad de este aporte de energía es radiación visible y la mitad es infrarrojo cercano. La salida de energía de la Tierra también es de aproximadamente 8.36 J/cm 2 -min, ocurriendo como infrarrojo de onda más larga. Debido a que las tasas de entrada y salida de energía son iguales, no hay almacenamiento neto de energía, y la temperatura media de la superficie del planeta se mantiene estable. Por lo tanto, como se señaló anteriormente, el presupuesto energético de la Tierra se puede caracterizar como un sistema de flujo continuo de suma cero.
Sin embargo, lo anterior no es exactamente cierto. A lo largo de escalas extremadamente largas de tiempo geológico, se ha producido una pequeña cantidad de almacenamiento de energía solar a través de una acumulación de biomasa no descompuesta que finalmente se transformó en combustibles fósiles. Además, se producen fluctuaciones relativamente menores a largo plazo en la temperatura superficial de la Tierra, lo que representa un elemento importante del cambio climático. Sin embargo, estas son excepciones cuantitativamente triviales a la afirmación de que la Tierra es un sistema de flujo continuo de suma cero para la energía solar.
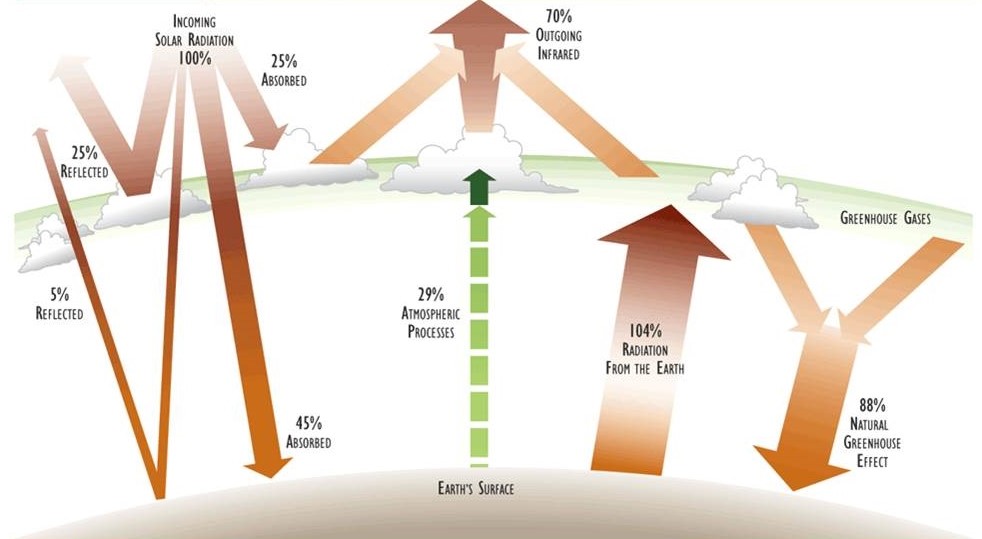
Aunque la cantidad de energía emitida por la Tierra eventualmente es igual a la cantidad de radiación solar que se absorbe, muchas transformaciones ecológicamente importantes ocurren entre la absorción inicial y la eventual re-radiación. Estos son los elementos internos del presupuesto de energía física del planeta (ver Figura 4.2). Los componentes más importantes se describen a continuación:
Reflexión — En promedio, la atmósfera y la superficie de la Tierra reflejan alrededor del 30% de la energía solar entrante de regreso al espacio exterior. La reflectividad de la Tierra (albedo) está influenciada por factores tales como el ángulo de la radiación solar entrante (que varía durante el día y a lo largo del año), las cantidades de cobertura de nubes reflectantes y partículas atmosféricas (también altamente variables), y el carácter de la superficie, especialmente los tipos y cantidades de agua (incluyendo nieve y hielo) y vegetación más oscura.
Absorción por la Atmósfera — Alrededor del 25% de la radiación solar incidente es absorbida por gases, vapores y partículas en la atmósfera, incluidas las nubes. La tasa de absorción es específica de la longitud de onda, siendo partes del rango infrarrojo intensamente absorbidas por los llamados gases “de efecto invernadero” (especialmente vapor de agua y dióxido de carbono; ver Capítulo 17). La energía absorbida se convierte en calor y se re-irradia como radiación infrarroja de una longitud de onda más larga que la inicialmente absorbida.
Absorción por la Superficie — En promedio, alrededor del 45% de la radiación solar entrante pasa a través de la atmósfera y es absorbida en la Tierra por materiales vivos y no vivos en la superficie, una trabsformación que aumenta su temperatura. Sin embargo, esta cifra de 45% es muy variable, dependiendo en parte de las condiciones atmosféricas, especialmente de la cobertura de nubes, y también de si la luz incidente ha pasado por el dosel de una planta. Si bien a largo plazo (años) e incluso a mediano plazo (días) el almacenamiento neto global de calor es esencialmente cero, en algunos lugares puede haber cambios sustanciales en el almacenamiento neto de energía térmica dentro del año. Esto ocurre en todas partes de Canadá por la estacionalidad de su clima, en que los ambientes son mucho más cálidos durante el verano que en el invierno. Sin embargo, casi toda la energía absorbida se disipa finalmente por la re-radiación de la superficie como infrarrojo de onda larga.
Evaporación del agua — Parte de la energía térmica de las superficies vivas y no vivas hace que el agua se evapore en un proceso conocido como evapotranspiración. Este proceso tiene dos componentes: la evaporación del agua de lagos, ríos, arroyos, rocas húmedas, suelo y otros sustratos no vivos, y la transpiración del agua de cualquier superficie viva, particularmente del follaje de las plantas, pero también de las superficies corporales húmedas y los pulmones de los animales.
Derretimiento de Nieve y Hielo — La energía térmica absorbida también puede hacer que el hielo y la nieve se derrita, lo que representa una transformación energética asociada con un cambio de estado del agua de una forma sólida a una líquida.
Corrientes de Viento y Agua — Existe una distribución muy desigual del contenido de energía térmica en y cerca de la superficie de la Tierra, siendo algunas regiones bastante frías (como el Ártico) y otras mucho más cálidas (los trópicos). Debido a esta asignación irregular de calor, la superficie desarrolla procesos para disminuir los gradientes de energía transportando masa alrededor del globo, como por los vientos y las corrientes oceánicas (ver también Capítulo 3).
Fijación Biológica — Una porción muy pequeña pero ecológicamente crítica de la radiación solar entrante (con un promedio global de menos de 0.02%) es absorbida por la clorofila en plantas y algas y utilizada para impulsar la fotosíntesis. Esta fijación biológica permite que parte de la energía solar se almacene temporalmente como energía potencial en bioquímicos, sirviendo así como base energética para la productividad ecológica y la vida en la Tierra.
Energía en Ecosistemas
Un presupuesto de energía ecológica se enfoca en la absorción de energía por los organismos fotosintéticos y la transferencia de esa energía fija a través de los niveles tróficos de los ecosistemas (“trófico” se refiere a los medios de nutrición orgánica). Los ecologistas clasifican a los organismos en función de las fuentes de energía que utilizan.
Los autótrofos son capaces de sintetizar sus complejos bioquímicos utilizando compuestos inorgánicos simples y una fuente externa de energía para impulsar el proceso. La gran mayoría son fotoautótrofos, que utilizan la luz solar como su fuente externa de energía. Los fotoautótrofos capturan la radiación solar utilizando pigmentos fotosintéticos, el más importante de los cuales es la clorofila. Las plantas verdes son los ejemplos más abundantes de fotoautótrofos, pero las algas y algunas bacterias también son fotoautótrofas.
Un número mucho menor de autótrofos son los quimioautótrofos, que aprovechan parte del contenido energético de ciertos químicos inorgánicos para impulsar un proceso llamado quimiosíntesis. La bacteria Thiobacillus thiooxidans, por ejemplo, oxida los minerales sulfurados a sulfato y utiliza parte de la energía liberada durante esta reacción para quimiosintetizar moléculas orgánicas.
Debido a que los autótrofos son la base biológica de la productividad ecológica, los ecologistas se refieren a ellos como productores primarios. La fijación total de la energía solar por parte de todos los productores primarios dentro de un ecosistema se conoce como producción primaria bruta (GPP). Los productores primarios utilizan parte de esta producción para su propia respiración (R), es decir, para las funciones fisiológicas necesarias para mantener su salud y crecer. La respiración es la oxidación metabólica de los bioquímicos, y requiere un suministro de oxígeno y libera dióxido de carbono y agua como productos de desecho. La producción primaria neta (PNP) se refiere a la fracción de GPP que queda después de que los productores primarios hayan utilizado algunos para su propia respiración. En otras palabras: PNP = GPP — R.
La energía fijada por los productores primarios es la base para la productividad de todos los demás organismos, conocidos como heterótrofos, cuyos heterótrofos dependen de otros organismos, vivos o muertos, para abastecer la energía que necesitan. Los heterótrofos animales que se alimentan de plantas se conocen como herbívoros (o consumidores primarios); tres ejemplos familiares son ciervos, gansos y saltamontes. Los heterótrofos que consumen otros animales se conocen como carnívoros (o consumidores secundarios), como el lobo maderero, el halcón peregrino, los tiburones y las arañas. Algunas especies se alimentan tanto de biomasa vegetal como animal y son conocidas como omnívoros, el oso pardo es un buen ejemplo, al igual que nuestra propia especie. Muchos otros heterótrofos se alimentan principalmente de materia orgánica muerta y se llaman descomponedores o detrívoros, como buitres, lombrices de tierra y la mayoría de hongos y bacterias.

La productividad es la producción expresada como una función de tasa, es decir, por unidad de tiempo y área. La productividad en los ecosistemas terrestres a menudo se expresa en unidades como kilogramos de biomasa seca (o su equivalente energético) por hectárea por año (kg/ha-y o kJ/ha-y), mientras que la productividad acuática a menudo se da como gramos por metro cúbico por año (g/m 3 -y).
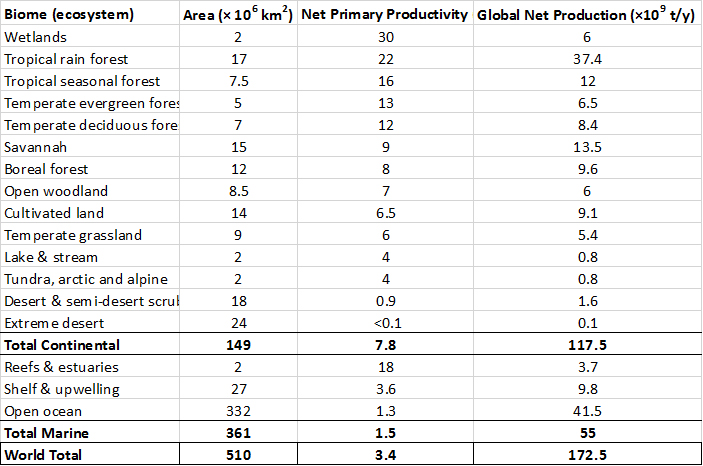
Se han realizado muchos estudios sobre la productividad de los diversos niveles tróficos en los ecosistemas. Por ejemplo, estudios de un bosque natural de encino-pino encontraron que la fijación total de la energía solar por la vegetación (la producción primaria bruta anual) fue equivalente a 4.81 × 104 kJ/m2-y (48 100 kJ/m 2 -y) (Odum, 1993). Esta tasa de fijación fue equivalente a menos de 0.1% de la entrada anual de radiación solar. Debido a que las plantas utilizaron 2.72 × 104 kJ/m 2 -y durante su respiración, la productividad primaria neta fue de 2.09 × 104 kJ/m 2 -y, representada principalmente por la biomasa creciente de los árboles. Los diversos organismos heterótrofos del bosque utilizaron 1.26 × 104 kJ/m 2 -y para apoyar su respiración. En última instancia, la acumulación neta de biomasa por todos los organismos del ecosistema (referida como la productividad neta del ecosistema) fue equivalente a 0.83 × 104 kJ/m 2 -y, o 8.3 × 103 kJ/m 2 -y.
Las productividades primarias de las principales clases de ecosistemas del mundo se resumen en el Cuadro 4.1. Tenga en cuenta que la tasa de producción es mayor en bosques tropicales, humedales, arrecifes coralinos y estuarios. La producción para cada tipo de ecosistema se calcula como su productividad multiplicada por su área. Sin embargo, las mayores cantidades de producción ocurren en los bosques tropicales y el océano abierto. Tenga en cuenta que el océano abierto tiene una productividad relativamente pequeña, pero su producción global es grande debido a su enorme área.
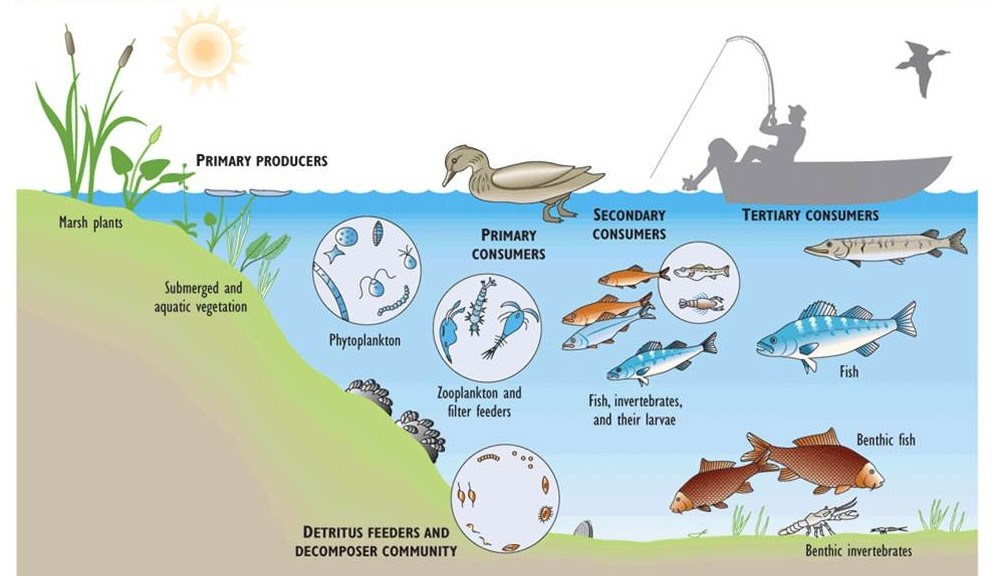
Una cadena alimentaria ecológica es un modelo lineal de relaciones de alimentación entre especies. Un ejemplo de una cadena alimentaria sencilla en el norte de Canadá son los líquenes y juncias, que son comidos por los caribú, que son comidos por los lobos. Una red alimentaria es un modelo más complejo de relaciones alimentarias, ya que describe las conexiones entre todas las cadenas alimentarias dentro de un ecosistema. Los lobos, por ejemplo, son depredadores oportunistas que pueden alimentarse de liebre con raquetas de nieve, topillos, lemming, castor, aves y otras presas además de su presa habitual de ciervos, alces y caribú. Por lo tanto, los lobos participan en diversas cadenas alimentarias dentro de su ecosistema. Sin embargo, ningún depredador natural se alimenta de lobos, a los que por lo tanto se les conoce como carnívoros superiores o depredadores superiores.
La Figura 4.3 ilustra elementos importantes de la red alimentaria del lago Erie, uno de los Grandes Lagos. En este gran lago, los ambientes de aguas poco profundas soportan las plantas acuáticas, mientras que el fitoplancton se presenta en toda la columna de agua superior. Las plantas de aguas poco profundas son consumidas por patos, ratas almizcleras y otros herbívoros, mientras que el fitoplancton es consumido por pequeños crustáceos (zooplancton) y filtro-alimentadores de fondo como las almejas. El zooplancton es comido por peces pequeños como el olor, que son consumidos por peces más grandes, que eventualmente pueden ser comidos por cormoranes, águilas calvas o humanos. La biomasa muerta de cualquier nivel de la red alimentaria puede asentarse hasta el fondo, donde ingresa a una red alimentaria detrítica y es consumida por animales pequeños y finalmente descompuesta por bacterias y hongos.
De acuerdo con la segunda ley de la termodinámica, la transferencia de energía en las redes alimentarias siempre es ineficiente porque parte de la energía fija debe ser convertida en calor. Por ejemplo, cuando un herbívoro consume biomasa vegetal, solo parte del contenido energético puede ser asimilado y transformado en su biomasa. El resto se excreta en las heces o se utiliza en la respiración (Figura 4.4). En consecuencia, en todos los ecosistemas la cantidad de productividad por parte de los autótrofos es siempre mucho mayor que la de los herbívoros, que a su vez siempre es mucho mayor que la de sus depredadores. Como una generalización amplia, hay alrededor de un 90% de pérdida de energía en cada etapa de transferencia. En otras palabras, la productividad de los herbívoros es solo alrededor del 10% de la de su alimento vegetal, y la productividad del primer nivel de carnívoros es solo del 10% de la de los herbívoros de los que se alimentan.
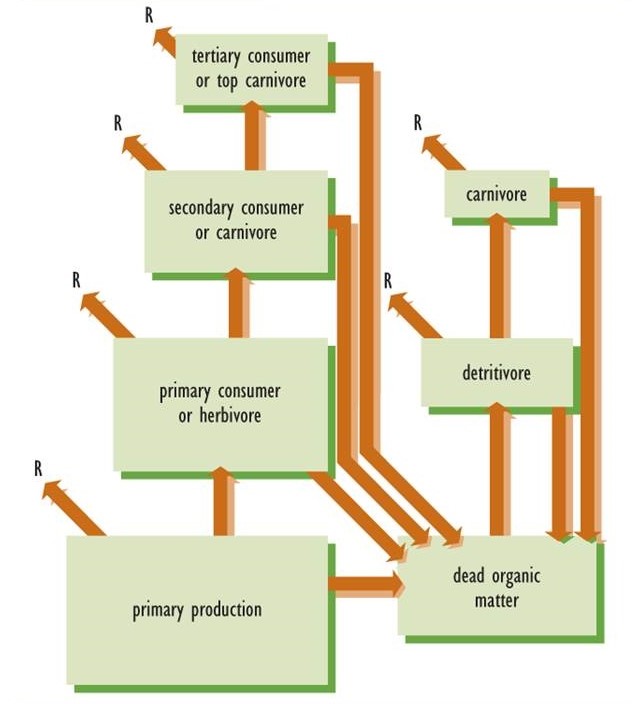
Estas relaciones de productividad se pueden mostrar gráficamente utilizando una llamada pirámide ecológica para representar la estructura trófica de un ecosistema. Las pirámides ecológicas se organizan con la productividad de las plantas en el fondo, la de los herbívoros por encima de las plantas y los carnívoros por encima de los herbívoros. Si el ecosistema sostiene a los carnívoros superiores, están representados en la cúspide de la pirámide. Los tamaños de las cajas tróficas en la Figura 4.4 sugieren la estructura piramidal de la productividad del ecosistema.
La segunda ley de la termodinámica se aplica a la productividad ecológica, función que está directamente relacionada con el flujo de energía. Sin embargo, la segunda ley no explica directamente la biomasa acumulada de un ecosistema. En consecuencia, es solo la estructura trófica de la productividad la que siempre tiene forma de pirámide. En algunos ecosistemas, otras variables pueden tener una estructura trófica piramidal, como las cantidades de biomasa (cultivo en pie) presentes en momentos específicos, o los tamaños o densidades de las poblaciones. Sin embargo, estas variables particulares no tienen forma piramidal en todos los ecosistemas.
Por ejemplo, en el océano abierto, el fitoplancton son los productores primarios, pero a menudo mantienen una biomasa similar a la del pequeño zooplancton que se alimenta de ellos. Las células del fitoplancton tienen una vida relativamente corta y su biomasa se da vuelta rápidamente debido a sus altas tasas de productividad y mortalidad. En contraste, los animales individuales del zooplancton tienen una vida más larga y mucho menos productivos que el fitoplancton. En consecuencia, la productividad del fitoplancton es mucho mayor que la del zooplancton, aunque en cualquier momento en particular estos niveles tróficos puedan tener una biomasa similar.
Algunos ecosistemas pueden incluso tener una pirámide invertida de biomasa, caracterizada por una biomasa menor de plantas que de herbívoros. Esto ocurre a veces en pastizales, en los que las plantas dominantes son especies herbáceas relativamente pequeñas que pueden ser bastante productivas pero que no mantienen una gran biomasa. En comparación, algunos de los herbívoros que se alimentan de las plantas son animales grandes, de larga vida, los cuales pueden mantener una biomasa total mayor que la vegetación. Algunos pastizales templados y tropicales tienen una pirámide de biomasa invertida, especialmente durante la estación seca cuando puede haber grandes poblaciones (y biomasa) de herbívoros de larga vida como antílope, bisonte, venado, elefante, gacela, hipopótamo o rinoceronte. Sin embargo, de acuerdo con la segunda ley de la termodinámica, la productividad anual (o a largo plazo) de las plantas en estos pastizales es siempre mucho mayor que la de los herbívoros.
Además, las densidades poblacionales de los animales no son necesariamente menores que las de las plantas que comen. Por ejemplo, los insectos son los herbívoros más importantes en muchos bosques y comúnmente mantienen grandes poblaciones. En contraste, el número de árboles es mucho menor, porque cada planta individual es grande y ocupa mucho espacio. Los bosques suelen mantener muchos más herbívoros que los árboles y otras plantas, por lo que la pirámide de números está invertida en forma. Como en todos los ecosistemas, sin embargo, la pirámide de productividad forestal es mucho más amplia en la parte inferior que en la parte superior.
Debido a la ineficiencia de la transferencia de energía entre niveles tróficos, existen límites energéticos para el número de carnívoros superiores (como águilas, orcas, tiburones y lobos) que pueden ser sostenidos por un ecosistema. Para sostener una población viable de depredadores superiores, debe haber una producción adecuadamente grande de presas que estos animales puedan explotar. Esta presa debe, a su vez, estar sustentada por una productividad de planta apropiadamente alta. Debido a estas limitaciones ecológicas, solo ecosistemas extremadamente productivos o muy extensos pueden soportar a los principales depredadores. De todos los ecosistemas terrestres de la Tierra, ninguno soporta más especies de carnívoros de orden superior que las sabanas y pastizales de África. Los más destacados de estos depredadores superiores son el guepardo, la hiena, el leopardo, el león y el perro salvaje. Esta riqueza inusualmente alta de depredadores superiores se puede sostener porque estos ecosistemas africanos son inmensos y bastante productivos de vegetación, excepto durante años de sequía. En contraste, la tundra del norte de Canadá puede soportar solo una especie natural de depredador superior, el lobo. Aunque la tundra es un bioma extenso, es un ecosistema relativamente improductivo.
Algunas poblaciones humanas preindustriales funcionaron como depredadores superiores. Esto incluyó a ciertos pueblos aborígenes de Canadá, como los inuit del Ártico y muchas culturas de las Primeras Naciones del bosque boreal. Como consecuencia ecológica de su estrategia de alimentación de orden superior dentro de su red alimentaria, estas culturas no pudieron mantener grandes poblaciones. En la mayoría de las economías modernas, sin embargo, los humanos interactúan con los ecosistemas de una manera omnívora: cosechamos una gama extremadamente amplia de alimentos y otros productos de biomasa de microbios, hongos, algas, plantas y animales invertebrados y vertebrados. Una de las consecuencias de este tipo de alimentación es que se puede sostener a una gran población humana.
Cuestiones Ambientales 4.1. Vegetarianismo y Eficiencia Energética
La mayoría de las personas tienen una dieta omnívora, es decir, comen una amplia variedad de alimentos tanto de origen vegetal como animal. Los vegetarianos, sin embargo, no comen carne u otros alimentos producidos al matar aves, peces, mamíferos u otros animales. Algunos vegetarianos, conocidos como veganos, no comen ningún alimento de origen animal, incluyendo queso, huevos, miel o leche. Las personas pueden optar por adoptar un estilo de vida vegetariano por diversas razones, entre ellas las que se centran en la ética de la crianza y sacrificio de animales y los beneficios para la salud de una dieta balanceada que no incluya productos animales. Además, existen grandes beneficios ambientales del vegetarianismo. Son cuotas para evitar ciertos contaminantes del aire, el agua y el suelo, y reducir la conversión del hábitat natural en agroecosistemas utilizados para la cría de ganado. Además, se necesita mucha menos energía para alimentar a una población de humanos vegetarianos que a los omnívoros.
Los animales cultivados comen una gran cantidad de comida. En la agricultura industrial practicada en los países desarrollados, incluido Canadá, la ganadería se cría principalmente con una dieta de productos vegetales, incluido el grano cultivado. Algunos vegetarianos argumentan que si ese grano fuera alimentado directamente a las personas, las cantidades totales de cereales y tierras agrícolas necesarias para apoyar a la población humana serían mucho menores. Este argumento se basa en la ineficiencia de la transferencia de energía entre niveles tróficos, que examinamos en este capítulo en un contexto más ecológico. Este argumento de eficiencia energética es muy convincente para los animales que se alimentan de granos y otros alimentos concentrados. Es menos relevante para el ganado que pasa toda o parte de su vida pastando en pastizales silvestres —en ese contexto ecológico, los animales rumiantes como vacas y ovejas están comiendo biomasa vegetal que los humanos no podrían consumir directamente y por lo tanto están produciendo alimentos que de otro modo no estarían disponibles.
De manera similar, muchos pollos, cerdos y otros animales son alimentados con desechos de alimentos (por ejemplo, de restaurantes) y subproductos de procesamiento (como sacrificios y peladuras de verduras y frutas, y puré de granos de cervecerías) que no son adecuados para el consumo humano. Se ha estimado que alrededor del 25% de las tierras de cultivo mundiales se está utilizando para cultivar granos y otros alimentos para el ganado, y que 37% de la producción mundial de cereales se alimenta a animales agrícolas. En Norteamérica, sin embargo, alrededor del 70% de la producción de granos se alimenta al ganado. Y hay un inmenso número de animales agrícolas: a nivel mundial, hay más de 3 mil millones de vacas, cabras y ovejas, y al menos 20 mil millones de pollos. Solo las vacas comen el equivalente a las necesidades calóricas de 8 a 9 mil millones de personas.
La eficiencia de asimilación es una medida del porcentaje del contenido energético de un alimento ingerido que es absorbido por el intestino y por lo tanto disponible para apoyar las necesidades metabólicas de un animal. Esta eficiencia varía entre grupos de animales y también depende del tipo de alimento que se consuma. Los animales herbívoros suelen tener una eficiencia de asimilación del 20 al 50%, siendo la tasa más pequeña para alimentos duros, fibrosos y de mala calidad como pasto y paja, y el más grande para alimentos de mayor calidad como el grano. Los carnívoros tienen una mayor eficiencia de asimilación, alrededor del 80%, debido a que su alimento es tan denso en proteínas y grasas. En general, se necesitan alrededor de 16 kg de alimento para producir 1 kg de carne de res en un engaradero. Las proporciones para otros animales son 6:1 para la carne de cerdo, 3:1 para el pollo y 2:1 a 3:1 para los peces de piscifactoría. Estas ineficiencias de asimilación se evitarían si la gente comía directamente el grano que consume el ganado.
La energía ecológica no es la única consideración en la eficiencia energética del vegetarianismo. También se utilizan enormes cantidades de energía para convertir los ecosistemas naturales en tierras de cultivo, para cultivar y administrar los agroecosistemas, para transportar productos básicos, procesar y empaquetar alimentos, y para transportar, tratar o eliminar los materiales de desecho. Estos gastos energéticos también se reducirían sustancialmente si más personas tuvieran una dieta y estilo de vida vegetarianos. Claramente, los vegetarianos tienen una menor “huella ecológica” asociada a sus hábitos alimenticios (ver Cuadro 25.1 en el Capítulo 25).
Conclusiones
La energía puede existir en varios estados, pero las transformaciones de una a otra deben obedecer las leyes de la termodinámica. Los organismos y ecosistemas se degradarían espontáneamente si no tuvieran acceso continuo a fuentes externas de energía. En última instancia, la luz solar es la fuente clave de energía que soporta casi toda la vida y los ecosistemas. La luz solar es utilizada por los fotoautótrofos para combinar dióxido de carbono y agua en moléculas orgánicas simples a través del proceso metabólico de la fotosíntesis. La energía fija de la biomasa vegetal soporta redes alimentarias ecológicas. Las plantas pueden ser consumidas por herbívoros y la energía obtenida se utiliza para apoyar su propio crecimiento. Los herbívoros pueden entonces ser comidos por los carnívoros. Biomasa muerta apoya una red alimentaria descompositora. La luz solar también impulsa importantes funciones planetarias, como los sistemas hidrológicos y climáticos. Las actividades humanas pueden tener una influencia grande y degradante en las redes alimentarias, e incluso en el sistema climático de la Tierra al influir en la intensidad del efecto invernadero del planeta.
Preguntas para revisión
- ¿Qué formas de energía se describen en este capítulo? ¿Cómo se puede cambiar cada uno en otras formas?
- ¿Cuáles son las leyes primera y segunda de la termodinámica? ¿Cómo gobiernan las transformaciones de la energía?
- ¿Cuáles son los principales elementos del presupuesto de energía física de la Tierra?
- ¿Por qué la estructura trófica de la productividad ecológica tiene forma de pirámide?
Preguntas para Discusión
- Según la segunda ley de la termodinámica, los sistemas siempre se mueven espontáneamente hacia una condición de mayor entropía. Sin embargo, la vida y los ecosistemas en la Tierra representan sistemas locales donde continuamente se genera entropía negativa. ¿Qué condiciones permiten que exista esta aparente paradoja?
- ¿Por qué no hay depredadores naturales de orden superior que maten y coman leones, lobos y tiburones?
- ¿Por qué sería más eficiente que la gente sea vegetariana? Discuta su respuesta en vista de la estructura piramidal de la productividad ecológica.
- Haz una lista de las fuentes clave y transformaciones de energía que te apoyan a ti y a tus actividades en un día típico. ¿Cuál es la fuente definitiva de cada uno de los recursos energéticos que utiliza (como la luz solar y los combustibles fósiles)?
Explorando problemas
- Como parte de un estudio del ciclo de contaminantes, se le ha pedido que describa la red alimentaria de dos ecosistemas locales. Uno de los ecosistemas es un bosque natural (o pradera) y el otro es un área utilizada para cultivar trigo (u otro cultivo). ¿Cómo determinaría los principales componentes de las redes alimentarias de estos ecosistemas, las especies que ocurren en sus niveles tróficos y las interacciones entre las diversas especies que están presentes?
Referencias citadas y lecturas adicionales
Botkin, D.B. y E.A. Keller. 2014. Ciencias Ambientales: La Tierra como Planeta Viviente. 9a ed. Wiley & Hijos, Nueva York, NUEVO.
Freedman, B. 1995. Ecología Ambiental. 2a ed. Prensa Académica, San Diego, CA. Gates, D.M. 1985. Energía y Ecología. Sinauer, Nueva York, NY.
Hinrichs, R.A. y M. Kleinbach. 2012. Energía: Su Uso y Medio Ambiente. 5ta ed. Brook Cole, Florencia, KY.
Houghton, J.T. 2009. Calentamiento Global. El informe completo, 4ta ed. Cambridge University Press, Cambridge, Reino Unido.
Odum, E.P. 1993. Ecología Básica. Saunders College Publishing, Nueva York, NY
Liu, P.I. 2009. Introducción a la Energía, Tecnología y Medio Ambiente. 2a ed. ASME Press, Nueva York, NY.
Sacerdote, J. 2012. Energía: Principios, Problemas, Alternativas. 8a ed. Kendall Hunt Publishing Co., Dubuque, IO.
Schneider, S.H. 1989. El clima cambiante. Científico Americano, 261 (3): 70-9
Whittaker, R.H. y G.E. Likens. 1975. La biosfera y el hombre. pp. 305-28. En: Productividad Primaria de la Biosfera. (H. Lieth y R.H. Whittaker, eds.). Springer-Verlag, Nueva York, NY.