
9: Ecología - De los individuos a la biosfera



- Última actualización
- 30 oct 2022
- Guardar como PDF

( \newcommand{\kernel}{\mathrm{null}\,}\)
Objetivos de aprendizaje
Después de completar este capítulo, podrás
- Describir cómo las especies se adaptan a diferentes niveles de estrés y perturbación en su hábitat.
- Explicar cómo ocurre el crecimiento poblacional, así como las limitaciones en el tamaño de la población.
- Enumerar los principales factores ambientales que influyen en las comunidades ecológicas.
- Describir qué se entiende por paisaje (y paisaje marino) y cómo las influencias ambientales afectan su dinámica espacial y temporal.
- Esbozar la hipótesis de Gaia y discutir su aplicabilidad al funcionamiento de la biosfera.
Introducción
La ecología es el estudio de las relaciones entre los organismos y su entorno. En el sentido que aquí se entiende, “ambiente” incluye tanto (a) factores no vivos, como temperatura, humedad, nutrientes y perturbaciones físicas, así como (b) organismos vivos, que ejercen influencias a través de la competencia, herbivoría, depredación y enfermedad, y proporcionando elementos de hábitat (como cuando los árboles proporcionar hábitat para especies que viven en un bosque). Debido a que todos los organismos y ecosistemas están sometidos a una multiplicidad de influencias, puede ser difícil predecir los efectos ecológicos de los cambios en las condiciones ambientales.
Algunas influencias ambientales son recursos que los organismos pueden explotar como oportunidades, lo que les permite obtener las necesidades de la vida y el sustento. Otras influencias ambientales son factores estresantes, o limitaciones en la productividad y el éxito reproductivo. Muchos factores estresantes operan de manera continua (crónica), como suele ser el caso de los factores climáticos, la contaminación del suelo y el agua, y muchas interacciones biológicas. Otros factores estresantes afectan a organismos y ecosistemas como eventos de perturbación, los cuales causan graves daños en un corto periodo de tiempo. A una perturbación le sigue un periodo prolongado de recuperación ecológica llamado sucesión. La perturbación puede ser causada por fuerzas naturales como un incendio forestal o tormenta de viento, o por influencias antropogénicas como la tala clara de un bosque o el arado de un campo.

Imagen 9.2. Esta población de alcatros norteños (Morus bassanus) se reproduce en Cape St. Mary's en Terranova. Fuente: B. Freedman. 
La ecología considera la estructura y función de la red de la vida en una jerarquía de niveles:
- Un organismo individual se define, en un contexto evolutivo, como una entidad genéticamente única. Sin embargo, algunas especies se propagan por mecanismos asexuales, y pueden desarrollar clones de “individuos” genéticamente idénticos.
- Una población es un grupo de individuos de la misma especie que coexisten en el tiempo y el espacio y que potencialmente pueden cruzarse entre sí.
- Una especie consiste en una o más poblaciones en las que los individuos pueden cruzarse potencialmente, y se aíslan reproductivamente de otros grupos similares.
- Una comunidad es un conjunto de poblaciones de diversas especies que coexisten e interactúan como una agrupación distintiva.
- Un ecopaisaje es una integración espacial de varios tipos de comunidades en un área extensa. Cada comunidad es un “parche” espacial y el ecsoscape comprende un mosaico dinámico, que se conoce como paisaje en ambientes terrestres y como paisaje marino en los marinos.
- La biosfera consiste en toda la vida y los ecosistemas en la Tierra y los ambientes donde ocurren.
Cada uno de estos niveles de ecología es significativo, y todos son relevantes para la ciencia ambiental. Sin embargo, estos diversos niveles de ecología no son totalmente discretos, todos están interconectados y cada nivel influye entre sí. Este capítulo examina cuestiones que son relevantes para los diversos niveles jerárquicos de la ecología.
Imagen 9.3. Los arrecifes de coral son ecosistemas de aguas poco profundas en mares tropicales, y son extremadamente ricos en especies, como lo ilustra esta comunidad cercana a Puerto Morelos, México. Fuente: A. Pinder

Individuos y especies
La autecología es el campo dentro de la ecología que se ocupa del estudio de individuos, poblaciones y especies. Los temas importantes en autecología incluyen los siguientes:
- diferencias entre especies en características de historia de vida y en adaptaciones a diversos tipos de condiciones ambientales
- influencias del medio ambiente en organismos individuales, incluidos los efectos sobre su desarrollo y comportamiento
- las causas de los cambios en el tamaño y composición de las poblaciones
Características de la historia de vida
Cada especie es única y puede ser descrita por sus atributos anatómicos, conductuales, bioquímicos y ecológicos. Estas características están determinadas en última instancia por la variación genética colectiva que existe entre los individuos que componen la especie.
Cada especie es única. Sin embargo, las especies pueden agregarse en grupos en función de las similitudes de sus atributos. Estas afinidades pueden deberse a la relación ancestral, debido a que especies relacionadas comparten aspectos de su historia evolutiva. Por ejemplo, todos los arces (género Acer) se parecen bastante y ocurren en hábitats de bosque templado. De igual manera, todos los miembros de la familia de los gatos (Felidae) tienen cierto parecido y son ecológicamente comparables en que todos son depredadores, aunque de diferentes presas y en diferentes tipos de hábitat. Sin embargo, las especies no relacionadas también pueden mostrar atributos similares, generalmente porque han tenido una historia de cambios análogos a través de un fenómeno conocido como convergencia evolutiva (o evolución paralela). La convergencia sugiere que, a través de la selección natural, especies no relacionadas que viven en entornos comparables pueden evolucionar para parecerse entre sí y desempeñar papeles funcionales similares en su ecosistema.
Existen muchos ejemplos de convergencia evolutiva entre grupos de organismos no relacionados. Por ejemplo, todas las plantas perennes (de larga vida) que crecen en ambientes áridos tienen la necesidad de conservar la humedad. Esta función crítica se ve potenciada por una forma de crecimiento que incluye adaptaciones para reducir la pérdida de agua, como un tronco cilíndrico y ramas, tejidos protegidos por una cutícula cerosa y sin hojas. La espinosidad es otro rasgo útil en un ambiente árido porque las espinas disuaden a los herbívoros de consumir biomasa y reservas de agua. Muchas plantas que habitan en el desierto han desarrollado una o más de estas adaptaciones, incluyendo especies de cactus (familia Cactaceae), euforbs (Euphorbiaceae) y suculentas (Crassulaceae). Aunque las especies de estas familias no están estrechamente relacionadas en un sentido evolutivo, pueden parecerse entre sí debido a la convergencia evolutiva.
También hay ejemplos de convergencia entre especies de animales. Una de ellas son las similitudes del lobo maderero (Canis lupus) de Eurasia y América del Norte y el lobo marsupial (tilacino, Thylacinus cynocephalus) de Australia. Otro ejemplo es la marmota (Marmota monax) de América del Norte y el wombat marsupial (Vombatus ursinus) de Australia. Además, los pingüinos (familia Spheniscidae) del hemisferio sur son similares a los araos, murres, frailecillos y alcas afines (familia Alcidae) del hemisferio norte.
Los ecologistas suelen categorizar las especies vegetales en función de su autecología. Un sistema se basa en las adaptaciones de las plantas para hacer frente a ciertos tipos de condiciones de hábitat. El ecologista Philip Grime ha sugerido que las estrategias vegetales pueden dividirse en tres categorías básicas, las cuales están determinadas por la historia de vida y su relación con el hábitat. Este sistema propone que dos grupos de factores ambientales —perturbación y estrés— tienen una fuerte influencia en la evolución de las estrategias de historia de vida vegetal. La alteración puede ser frecuente o infrecuente, y su intensidad es grave o leve. El estrés es una condición de sitio a largo plazo, y puede ser intenso si se asocia con una escasez extrema de humedad, luz o nutrientes, o inocuo si estos factores vitales están bien disponibles. Cualquier ambiente particular puede caracterizarse por la importancia de estos dos grupos de factores, lo que da como resultado cuatro tipos básicos de condiciones de hábitat:
- bajo estrés y perturbaciones raras
- bajo estrés pero perturbación frecuente
- estrés intenso pero perturbación rara
- estrés intenso y perturbación frecuente
Sin embargo, Grime sugiere que las plantas presentan solo tres estrategias primarias de historia de vida, ya que no pueden hacer frente a un ambiente estresante y frecuentemente perturbado (4 arriba). Las tres estrategias primarias de historia de vida son:
- Las plantas competidoras son dominantes en hábitats en los que las perturbaciones son raras y las tensiones ambientales son relativamente poco importantes. En tales condiciones, la competencia es la mayor influencia selectiva en la evolución de las plantas y en la organización de sus comunidades. Las plantas competitivas son efectivas para adquirir recursos y utilizarlos para lograr una posición dominante en su comunidad al interferir con la productividad de otras plantas. Las adaptaciones útiles en los competidores incluyen un crecimiento alto rápido, un dosel extendido y un sistema radicular ampliamente extendido, estos personajes ayudan a ocupar espacio y aprovechar los recursos. Además, las plántulas de muchas plantas competitivas pueden establecerse bajo un dosel cerrado.
- Los ruderales ocurren en ambientes frecuentemente perturbados con abundantes recursos, por lo que el estrés no es grande. Por lo tanto, las plantas ruderales están bien adaptadas para utilizar hábitats ricos pero temporales. Suelen ser efímeros e intolerantes al estrés y la competencia. Los ruderales producen grandes cantidades de semillas, las cuales suelen tener mecanismos de dispersión a larga distancia para que los hábitats recién alterados puedan ser colonizados.
- Los tolerantes al estrés se adaptan a ambientes marginales en términos de clima, humedad o suministro de nutrientes, pero con poca frecuencia perturbados y, por lo tanto, estables. Son típicos de ambientes árticos, desérticos y otros estresantes, y generalmente son cortos, de crecimiento lento e intolerantes a la competencia.
Otro sistema de categorización de organismos, más comúnmente aplicado a los animales, involucra dos grupos de características de historia de vida. Uno consiste en organismos de vida más larga que producen relativamente poca progenie, pero invierten muchos recursos en cada uno para mejorar sus posibilidades de supervivencia. Estas son conocidas como especies seleccionadas por K. El otro grupo, denominado r-seleccionado, incluye especies de corta vida que producen grandes cantidades de crías pequeñas, cada una de las cuales tiene una probabilidad relativamente pequeña de supervivencia, pero debido a las enormes cantidades es probable que algunas persista. Las especies seleccionadas K son dominantes en hábitats maduros relativamente estables en los que la competencia es la influencia controladora en la estructura comunitaria, mientras que las especies seleccionadas por r ocurren en hábitats más jóvenes recientemente perturbados en los que los recursos están disponibles gratuitamente y es posible un rápido crecimiento poblacional. (La fuente de las etiquetas “K” y “r” proviene de la ecuación logística, elemento fundamental de la ecología poblacional que, por simplicidad, no se examina aquí).
Las especies también pueden ser consideradas en términos de otros aspectos de su estrategia reproductiva, como la frecuencia con la que se reproducen. Algunas especies solo tienen un evento reproductivo durante su vida, generalmente muriendo después. Este tipo de reproducción, conocida como semelpar, se observa en plantas anuales y bienales, muchos insectos y otros invertebrados, y salmón del Pacífico. La mayoría de las especies semelparas son de corta duración, pero algunas pueden vivir muchos años, acumulando gradualmente suficiente energía para sostener un esfuerzo reproductivo masivo y “big-bang”. La reproducción semelpar es favorecida en hábitats ricos que frecuentemente se alteran, y es común entre especies ruderales y seleccionadas por r.
Las especies que se reproducen varias veces durante su vida son conocidas como iteróparas. Se trata típicamente de especies de larga vida que viven en hábitats estables. Las especies Iteróparas pueden producir un gran número de crías pequeñas (seleccionadas por r), o pueden producir menos jóvenes más grandes, cada una de las cuales recibe una inversión sustancial de recursos parentales (seleccionados K).
Individuos y Medio Ambiente
La autecología también aborda la vida de los organismos individuales y cómo son influenciados por sus entornos físicos y biológicos.
Como examinamos en el Capítulo 6, todos los organismos individuales tienen un complemento fijo de información genética, conocido como su genotipo. Sin embargo, la expresión de información genética (el fenotipo) está influenciada por las condiciones ambientales, fenómeno conocido como plasticidad fenotípica. Si los individuos experimentan condiciones ambientales difíciles, la expresión fenotípica de su potencial genético puede incluir una tasa de crecimiento subóptima y la producción de poca o ninguna progenie. En contraste, otros individuos que viven en un ambiente más benigno pueden lograr una mayor productividad y tener muchas crías. Esta última circunstancia, más prolífica, es altamente deseable en términos de que un individuo logre el “éxito” evolutivo. Por definición, los individuos exitosos han logrado maximizar su condición física, su contribución genética a las generaciones futuras.
El éxito de un organismo individual también se ve afectado por alteraciones impredecibles, que pueden resultar en lesiones o muerte prematura. Incluso si vive en un ambiente benigno, con buen acceso a las necesidades de la vida, un individuo desafortunado puede simplemente ser quemado por un incendio forestal, devorado por un depredador, debilitado por una enfermedad o atropellado por un camión.
Ecología de Población
El estudio de las poblaciones es otro aspecto de la autecología. La abundancia de todas las especies cambia con el tiempo en respuesta a factores ambientales que afectan a cuatro variables relacionadas con la población (o demográficas): tasa de natalidad (BR), tasa de inmigración (IR), tasa de mortalidad (DR) y tasa de emigración (ER). El cambio en el tamaño de la población (ΔP) durante una unidad de tiempo (digamos, un año) se describe utilizando la siguiente ecuación:
ΔP = BR — DR + IR — ER
Esta relación demográfica es cierta para todas las especies, incluidos los humanos. En algunos casos, las poblaciones aisladas (o cerradas) no reciben ninguna inmigración de nuevos individuos y no pierden ninguna ante la emigración. En tales condiciones, ΔP se calcula como BR — DR, un valor conocido como la tasa natural de cambio poblacional.
A menudo, ΔP se expresa como un cambio porcentual al dividir su valor por el tamaño inicial de la población; por ejemplo, una población de 100 individuos que aumenta en 10 en un año tiene una tasa de crecimiento anual del 10%. Si el cambio porcentual en una población es constante a lo largo del tiempo, habrá una tasa acelerada de incremento o disminución, llamada cambio exponencial.
Imagínese una circunstancia en la que una pareja fértil de individuos logra descubrir un nuevo hábitat adecuado, uno que no ha sido ocupado previamente por su especie. En tales condiciones, los individuos fundadores se criarán y la población crecerá con el tiempo. Inicialmente, los recursos son abundantes y no limitan el crecimiento de la población. Durante ese periodo la tasa porcentual de incremento será constante, estando limitada sólo por la rapidez con la que se pueda producir la progenie (la tasa de natalidad) y llegar a ser fértiles (la tasa de maduración), y contrarrestada únicamente por las muertes que pudieran ocurrir. Esta es la tasa máxima de crecimiento poblacional, la cual está limitada únicamente por la biología de la especie y no por la competencia por los recursos, y se le conoce como la tasa intrínseca de incremento poblacional. Cualquier población que esté creciendo a la tasa intrínseca de incremento (o de hecho a cualquier tasa porcentual fija) explotará rápidamente en abundancia (ver En Detalle 9.1).
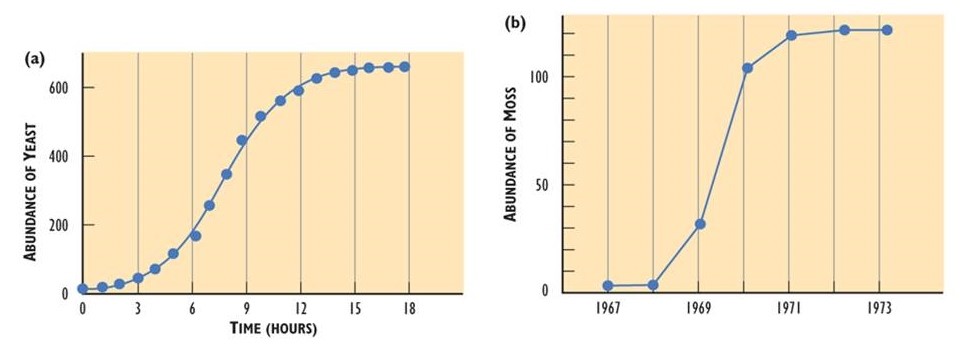
Eventualmente, sin embargo, la población se acercará a la capacidad de carga del hábitat disponible, o a la población que pueda ser sustentada sin causar que los recursos se vuelvan limitantes, u otros daños ambientales. En o más allá de la capacidad de carga, las oportunidades se ven limitadas por la limitada disponibilidad de recursos, por lo que los individuos de la población deben competir entre sí. La competencia intensa da como resultado estrés fisiológico, lo que puede provocar una disminución en la tasa de natalidad y un aumento en la tasa de mortalidad. En algunos casos, la tasa de incremento poblacional puede entonces disminuir a cero, lo que ocurre cuando la tasa de natalidad es igual a la tasa de mortalidad. Esta condición se conoce como crecimiento poblacional cero (ZPG). Si se mantiene ZPG, el tamaño de la población eventualmente se nivelará, tal vez a un nivel apropiado a la capacidad de carga del hábitat. Una curva poblacional de este tipo se conoce como “en forma de S”.
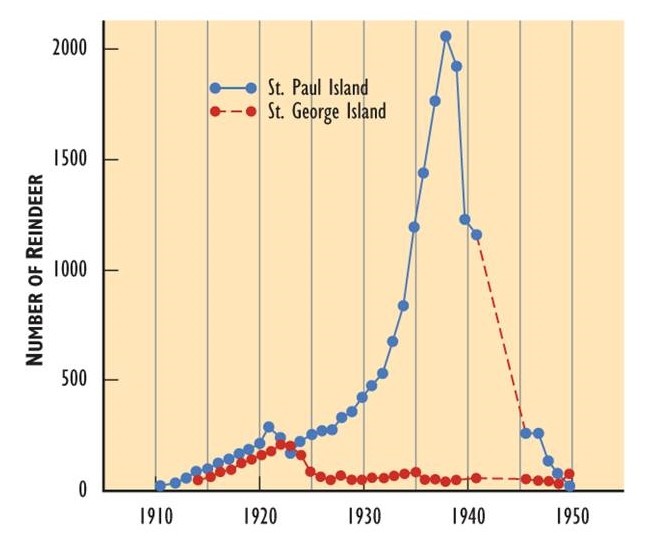
Sin embargo, el crecimiento exponencial anterior puede haber resultado en una abundancia que superó lo que el hábitat podría soportar. Tal sobrepoblación degradaría el ambiente, resultando en una disminución de su capacidad de carga. Si esto sucede, el tamaño de la población disminuirá a través de un incremento en la tasa de mortalidad, o tal vez por una oleada de emigración en busca de nuevos hábitats. Estos pueden resultar en una oscilación de la abundancia alrededor de la capacidad de carga, o en una caída rápida en el número de individuos en la población. Por lo general, un choque lleva a la población a un nivel por debajo de la capacidad de carga, creando una circunstancia para un crecimiento demográfico renovado. En hábitats pequeños, sin embargo, el choque puede ser lo suficientemente masivo como para extirpar a una población local.
Los ecologistas poblacionales han desarrollado modelos matemáticos de dinámica poblacional que dan cuenta de las influencias de factores tales como la tasa intrínseca de incremento poblacional, la capacidad de carga de los hábitats, los efectos de la depredación y la enfermedad, e incluso los efectos de perturbaciones impredecibles. Estos modelos se describen en libros de texto introductorios de ecología y no se tratan aquí con ningún detalle. Para los presentes propósitos, hay varios puntos importantes a entender sobre la ecología poblacional:
- Las poblaciones de todas las especies son dinámicas. Cambian con el tiempo debido a las diferentes tasas de nacimiento, muerte, inmigración y emigración.
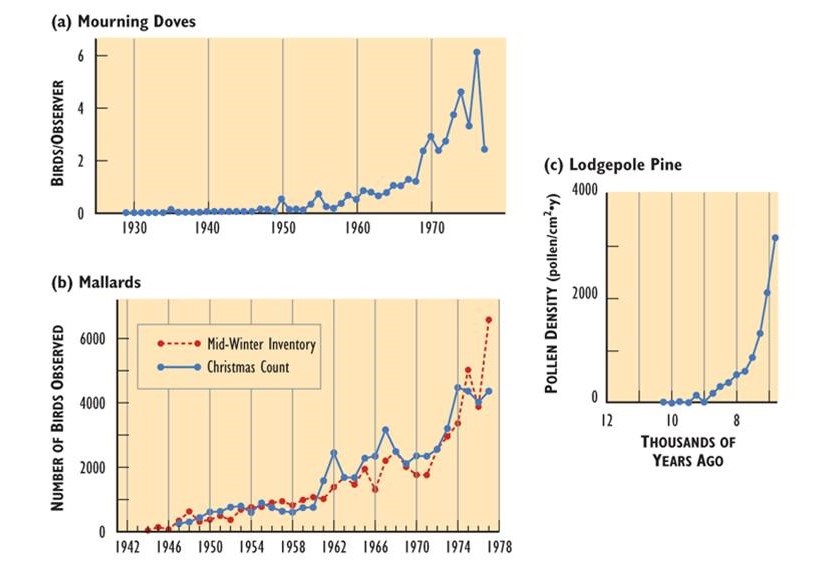
- Las poblaciones de todas las especies pueden, potencialmente, aumentar rápidamente en condiciones en las que la disponibilidad de recursos y otros factores no son limitantes. En la Figura 9.1 se ilustran ejemplos de rápido crecimiento poblacional. Sin embargo, el crecimiento ilimitado no puede sostenerse; en todos los casos de la Figura 9.1, el tamaño de la población finalmente se estabilizó, disminuyó o se estrelló.
- En última instancia, la abundancia sustentable de una especie está limitada por la capacidad de carga del hábitat disponible. Ejemplos de crecimiento poblacional que se nivelan en la capacidad de carga del hábitat se ilustran en la Figura 9.2.
- Algunas poblaciones son relativamente estables. Por lo general, existen en entornos en los que la disponibilidad de recursos es predecible para que se pueda lograr un equilibrio con la capacidad de carga. Por ejemplo, se producen relativamente pocos cambios en las poblaciones anuales de árboles que crecen en bosques antiguos, a menos que se produzca una perturbación rara y catastrófica.
- Otras poblaciones son relativamente dinámicas, cambiando mucho con el tiempo y raramente logrando incluso un equilibrio a corto plazo con la capacidad de carga de su hábitat. Esto es comúnmente cierto en especies que viven en hábitats que son perturbados frecuentemente o se encuentran en una etapa temprana y relativamente dinámica de sucesión. Algunas poblaciones son cíclicas, logrando grandes abundancias a intervalos regulares, intercaladas por periodos más largos de menor abundancia. Las poblaciones cíclicas son obviamente inestables a corto plazo, pero pueden ser estables a largo plazo.
- Las poblaciones que superan la capacidad de carga de su hábitat nunca son sustentables en ese alto nivel, debido al daño ambiental que se ocasiona. Las poblaciones insostenibles eventualmente se estrellan a una menor abundancia y a veces a la extinción. La Figura 9.3 muestra un ejemplo de rápido crecimiento poblacional que resultó en la degradación del hábitat y un posterior desplome poblacional. Las poblaciones también pueden chocar por otras razones, como la ocurrencia repentina de una enfermedad mortal. Esto sucede con el olmo blanco nativo (Ulmus americana) de América del Norte, que está siendo diezmado por un patógeno introducido (el hongo de la enfermedad del olmo holandés, Ceratocystis ulmi) al que este árbol tiene poca inmunidad. Otras causas de choques poblacionales incluyen niveles insostenibles de depredación y disturbios extensos como incendios forestales o despejados.
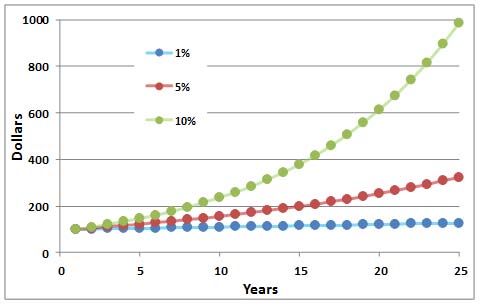
En Detalle 9.1. Crecimiento Exponencial
Una tasa constante de incremento conduce a un rápido crecimiento en el tamaño de una población. Esto sucede por la misma razón que el dinero invertido a una tasa de interés fija aumentará rápidamente en cantidad. Este fenómeno, conocido en las finanzas como interés compuesto, se ilustra a continuación.
Consideremos, por ejemplo, una inversión de $100 realizada a una tasa de interés fija del 10% anual, bloqueada por un periodo de 10 años. Después del Año 1, el depósito inicial crece a 110 dólares, lo que representa la inversión inicial más los intereses acumulados. En el Año 2, la tasa de interés del 10% se aplica a los $110, por lo que los intereses devengados son mayores ($11) que en el Año 1 ($10). En el Año 3, el interés del 10% se aplica a los 121 dólares acumulados, por lo que el interés devengado es aún mayor ($12.10), y el valor devengado de la inversión es de $133.10. Al finalizar el año 4, la inversión inicial de $100 vale $146.41. Es entonces $161.05 al final del Año 5... y $259.37 al final del Año 10, lo que representa un impresionante retorno del 159% sobre la inversión inicial. Claramente, una tasa de interés compuesta conduce a un rápido aumento del capital.
El crecimiento exponencial se refiere al crecimiento acelerado de una cantidad inicial debido a una tasa constante de incremento. A veces se calcula un parámetro importante conocido como el tiempo de duplicación, es el tiempo requerido para un doble aumento del capital. El tiempo de duplicación se puede calcular aproximadamente como 70 dividido por la tasa constante de aumento. En el ejemplo anterior, 70 dividido por 10% anual rinde siete. Por lo tanto, los 100 dólares iniciales se duplicarían en cantidad en sólo siete años, y los 200 dólares acumulados volverían a duplicarse (a 400 dólares) en otros siete años, y así sucesivamente siempre y cuando las condiciones de inversión no cambien.
Las matemáticas de interés compuesto también se pueden aplicar al crecimiento exponencial de poblaciones de organismos. Un ejemplo bastará: En 2015, la población humana global era de unos 7.3 mil 300 millones de personas, creciendo en alrededor de 1.3% anual. Por lo tanto, en sólo 54 años (es decir, 70 divididos por 1.3% anual), la población humana podría duplicarse a más de 14 mil millones, si la tasa de crecimiento no cambiara. Las implicaciones ambientales de tal incremento poblacional son inmensas (ver Capítulo 10).
Ecología Comunitaria
Una comunidad ecológica es una agregación de poblaciones que ocurren en el mismo tiempo y lugar como una agrupación distintiva, y que interactúan física, químicamente y/o conductual. El estudio de las relaciones entre especies dentro de las comunidades se conoce como sinecología. Estrictamente hablando, una comunidad consiste en todas las poblaciones vegetales, animales y microbianas que ocurren juntas en un sitio.
El nicho
Cada especie dentro de una comunidad explota el medio ambiente e interactúa con otras especies de una manera particular. Los ecologistas utilizan la palabra nicho para describir el papel de una especie en su comunidad, que también puede ser vista como su “ocupación” o sustento. Algunos nichos son relativamente estrechos y especializados, al igual que el nicho de murciélagos que se alimentan únicamente de insectos voladores de cierto tamaño, o avispas que polinizan solo una o unas pocas especies de plantas. Otros nichos, sin embargo, son mucho más amplios, como los de osos y humanos, los cuales se alimentan en un rango extremadamente amplio y afectan su ecosistema de diversas maneras.
El llamado nicho fundamental está determinado por el rango de tolerancia de una especie a las condiciones ambientales. Estas tolerancias se reflejan en las formas en que una especie obtiene su nutrición y cómo interactúa con otras especies, y están mediadas por aspectos de comportamiento, morfología y fisiología. En comparación, el nicho realizado refleja la gama de condiciones ambientales que una especie realmente logra explotar en la naturaleza. El nicho realizado es más pequeño que el nicho fundamental porque todas las especies están en cierta medida limitadas por interacciones biológicas como competencia, depredación y enfermedad.
Comunidades Funcionales
Debido a su complejidad, comunidades enteras rara vez son examinadas por ecologistas. Los estudios ecológicos suelen estar limitados por las cantidades de financiamiento y la amplitud de conocimientos disponibles. En cambio, la investigación a nivel comunitario generalmente implica el examen de grupos seleccionados de organismos similares, como “comunidades” de insectos, peces, aves, plantas o microbios. Aunque el alcance de este trabajo es limitado, sí permite a los ecologistas investigar aspectos importantes de la ecología comunitaria.
Las comunidades forestales, por ejemplo, contienen una amplia gama de organismos de diversas especies y tamaños, incluyendo plantas, animales y microorganismos. Las poblaciones de las diversas especies interactúan de múltiples maneras. Los árboles, por ejemplo, proporcionan la estructura física del hábitat, ponen alimentos a disposición de los herbívoros y dejan caer hojarasca que se descompone por especies de la red alimentaria detrítica. Otras interacciones dentro de una comunidad forestal incluyen depredación, parasitismo y enfermedades, así como simbiosis como polinización, dispersión de semillas y micorrizas radiculares. Debido a la complejidad inherente de las comunidades forestales, la mayoría de los estudios ecológicos solo investigan componentes seleccionados.
Este enfoque pragmático de la investigación a nivel comunitario puede ser ilustrado por estudios sobre los efectos ecológicos de la silvicultura realizados en los Marítimos por el autor y varios estudiantes. Para realizar este trabajo dividimos a la comunidad más grande en los siguientes grupos funcionales:
- árboles, los cuales definimos como plantas leñosas con un diámetro mayor a 10 cm
- arbustos, con un diámetro inferior a 10 cm pero mayores de 1 m, incluyendo individuos jóvenes del tamaño arbustivo de especies arbóreas así como arbustos “verdaderos”
- vegetación del suelo, incluyendo todas las plantas, musgos y líquenes, que crecen dentro de 1 m del suelo
- epífitas que crecen en otras plantas, como líquenes y musgos en las superficies cubiertas de corteza de los árboles
- pequeños mamíferos como ratones, musarañas, ratones de campo y ardillas
- mamíferos grandes como ciervos, osos y coyotes
- aves
- reptiles y anfibios
- insectos
- hongos y otros microorganismos en el suelo
Durante algunos de los estudios de aves, se realizó un trabajo específico con especies que anidan en cavidades en árboles. Estos comprenden un elemento “que requiere cavidades” de la comunidad aviar más grande. De igual manera, el trabajo sobre insectos y otros invertebrados ha involucrado grupos funcionales que viven en el suelo, en la madera muerta podrida o en el follaje. Pero incluso con todos estos (y otros) grupos funcionales, no logramos examinar todos los elementos de las comunidades forestales que estábamos estudiando.
Factores que influyen en las comunidades Las comunidades ecológicas se ven afectadas por diversos factores ambientales, particularmente los que se describen a continuación.
Especies Presentes — Obviamente, solo aquellas especies que están presentes en un hábitat, o son capaces de dispersarse en él, pueden desempeñar un papel en la comunidad que se desarrolla. La capacidad de una especie para colonizar un hábitat disponible está influenciada por su biología, interviniendo barreras como una cordillera u océano, el régimen de perturbación y otros factores. Cada vez más, los humanos están influenciando la composición de especies de las comunidades, a menudo introduciendo especies no indígenas más allá de su área de distribución natural.
Hábitat apropiado — Si un hábitat no es adecuado, entonces una especie en particular no podrá usarlo aunque sea capaz de dispersarse al sitio. Hay muchos aspectos de la idoneidad del hábitat, y todos ellos deben satisfacerse dentro de los límites de tolerancia de una especie para que se convierta en un componente de una comunidad.
Interacciones Biológicas — Las especies interactúan a través de herbivoría, depredación, competencia, enfermedad y simbiosis, incluyendo este último mutualismo, comensalismo y parasitismo. Todas estas interacciones pueden influir en la presencia y abundancia de especies dentro de las comunidades. Los siguientes ejemplos ilustran estas influencias.
Herbivoría: ocurre cuando los animales se alimentan de biomasa vegetal. Las larvas del looper de cicuta (Lambdina fiscellaria) son alimentadoras voraces en el follaje de abeto, abeto y otros árboles coníferos. Cuando las condiciones son adecuadas, esta polilla puede proliferar rápidamente, causando daños en una gran área de bosque, como sucede periódicamente en el este de Canadá. Los rodales defoliados desde hace varios años tienen muchos árboles muertos, lo que representa un elemento importante del cambio comunitario. La pérdida de gran parte del dosel forestal tiene efectos indirectos, como permitir que las plantas del sotobosque crezcan más vigorosamente. Los cambios en la vegetación afectan el hábitat disponible para especies de insectos, aves y otros animales. Los microorganismos y otros detritivores también se ven afectados debido a que se dispone de grandes cantidades de biomasa de árboles muertos para descomponerse.
Depredación: implica matar a un animal y comer a otro animal. Los depredadores pueden reducir en gran medida la abundancia de sus presas, cambiando así la estructura de la comunidad. Por ejemplo, durante el verano, la mayoría de las aves del bosque se alimentan de insectos, arañas y otros invertebrados, que son alimento nutritivo tanto para los adultos como para sus polluelos de rápido crecimiento. La depredación aviar puede cambiar la comunidad de invertebrados, como lo han demostrado estudios en los que pequeñas áreas de bosque fueron encerradas con redes. Esto excluía a los depredadores aviares, pero los invertebrados podían entrar o salir. En estas condiciones, la abundancia de muchos insectos y arañas aumentó, siendo las especies vulnerables a la depredación aviar las que más beneficiaron.

La competencia — ocurre cuando la demanda biológica de un recurso ecológico excede la oferta, provocando que los organismos interfieran entre sí. Las plantas, por ejemplo, a menudo compiten por el acceso a suministros limitados de luz solar, agua, nutrientes y espacio. Los animales pueden competir por alimentos, sitios de anidación, compañeros y otros recursos. La competencia intraespecífica ocurre cuando individuos de la misma especie compiten por el acceso a los recursos, mientras que la competencia interespecífica ocurre entre especies. Si una especie es particularmente efectiva en la cooptación de recursos en su propio beneficio, puede desplazar a otras especies, fenómeno conocido como desplazamiento competitivo (o en casos extremos, exclusión competitiva). Esto afecta la presencia y abundancia relativa de especies en la comunidad. Por ejemplo, el arce azucarero (Acer saccharum) es un árbol altamente competitivo en los bosques de frondosas del este de Canadá. Donde las condiciones ambientales son muy adecuadas para esta especie, puede dominar rodales maduros. Si se retiran grandes arces azucareros de un rodal, tal vez mediante una cosecha selectiva de madera, otras especies arbóreas (así como pequeños arces azucareros) se beneficiarán de la competencia reducida y crecerán más vigorosamente.
Enfermedad — es una relación patológica en la que la salud de plantas o animales sufre una infestación de otra especie, generalmente un microbio. Las enfermedades virulentas pueden provocar enormes cambios en la composición de las comunidades ecológicas. A principios del siglo XX, el castaño americano (Castanea dentata) fue afectado por el tizón del castaño (Endothia parasitica), un patógeno fúngico introducido. Debido a que las castañas tienen poca inmunidad a esta enfermedad, la especie fue prácticamente eliminada de los bosques del este de América del Norte en la década de 1950. Este cambio liberó a otras especies arbóreas de la competencia con el castaño previamente dominante, y rápidamente llenaron huecos en el dosel creado por su desaparición.
Simbiosis — se refiere a las relaciones íntimas que pueden ocurrir entre especies. Esto puede implicar una relación obligada en la que los simbiontes no pueden vivir separados, pero más comúnmente la asociación es algo flexible. Las simbiosis pueden influir en gran medida en el desempeño de las especies en entornos particulares al mejorar su capacidad competitiva y disminuir su vulnerabilidad a la depredación, enfermedad u otros estreses.
Los principales tipos de simbiosis son el mutualismo, en el que ambos socios se benefician; el parasitismo, en el que un organismo se beneficia y el otro se ve perjudicado; y el comensalismo, en el que un organismo se beneficia sin dañar al otro. Si bien las simbiosis son críticas para uno o ambos socios, también pueden afectar indirectamente el hábitat y los recursos disponibles para otros miembros de la comunidad.
Los líquenes son un ejemplo familiar de un mutualismo. Son una asociación obligada entre un hongo y una alga o una bacteria azul-verde. El hongo se beneficia de la productividad del socio fotosintético, mientras que este último obtiene un microhábitat relativamente húmedo y mejora el acceso a los nutrientes inorgánicos.
Otro mutualismo, llamado micorriza, es una asociación íntima entre los hongos y las raíces de las plantas vasculares. La planta se beneficia a través de un mayor acceso a nutrientes, especialmente fosfato, mientras que el hongo recibe exudados nutritivos de las raíces. Este mutualismo también proporciona un beneficio amplio a nivel comunitario a través del aumento de la productividad primaria. Muchas especies de leguminosas viven en un mutualismo con la bacteria Rhizobium japonicum, que fija el gas nitrógeno (N 2) en amoníaco, un nutriente crítico.
Otro mutualismo involucra especies de dinoflagelados (algas unicelulares) que viven dentro de los corales (pequeños animales coloniales), donde reciben protección y acceso a nutrientes. Los corales se benefician a través del acceso a la productividad fotosintética de las algas.
Muchos animales comen biomasa vegetal, pero pocos son capaces, por sí mismos, de digerir bioquímicos poliméricos complejos como la celulosa y la lignina. En consecuencia, muchos herbívoros viven en un mutualismo con microorganismos, que habitan en su intestino y secretan enzimas que digieren la celulosa y la lignina, poniendo a disposición del animal esas abundantes fuentes de nutrición. Vacas, venados y ovejas albergan sus microorganismos que ayudan a la digestión en una bolsa especializada de su proestómago, llamada rumen. Los humanos también albergan una comunidad diversa de microorganismos en su intestino, muchos de los cuales son importantes para nuestra nutrición.
Otros mutualismos incluyen las muchas especies de flores que son polinizadas por tipos particulares de insectos. La polinización es crucial para el éxito reproductivo de las plantas, mientras que los insectos se benefician de una abundante fuente alimenticia de néctar o polen. Además, los herbívoros de la comunidad se benefician de los frutos que se producen por la polinización, y a su vez pueden ayudar a dispersar las semillas de las plantas.
Un ejemplo de comensalismo es la comunidad epífita de plantas, líquenes y musgos que a menudo crecen en árboles grandes. Las epífitas obtienen un beneficio de la relación a través de un mayor acceso a la luz solar, pero los árboles hospedadores no se ven afectados en ningún grado significativo. Hay muchos ejemplos familiares de parasitismo, incluyendo pulgas en un perro y tenias en humanos. El parásito se beneficia al tomar la nutrición del huésped, pero el hospedador suele sufrir, e incluso puede morir por una infestación severa.

La perturbación es un evento de destrucción de alguna parte de una comunidad, un hecho que es seguido por un período a veces fraguado de recuperación ecológica llamado sucesión. Todas las comunidades son dinámicas, cambiando con el tiempo en su composición de especies y atributos funcionales (como productividad, descomposición y ciclo de nutrientes). Sin embargo, la tasa de cambio depende de la estabilidad de las condiciones ambientales, que es mayor en comunidades cercanas al punto final de una sucesión. En contraste, las comunidades más dinámicas están asociadas con las etapas más jóvenes de sucesión. Las perturbaciones pueden ocurrir en dos escalas espaciales.
- Las perturbaciones que reemplazan los estadios son causadas por incendios forestales, una epidemia de enfermedades, claros y otros eventos cataclísmicos. Este tipo de perturbación es extensa y da como resultado el reemplazo inmediato de una comunidad por otra diferente, seguida de un periodo de recuperación sucesional. Con el tiempo, la sucesión puede regenerar una comunidad similar a la que existía antes de la interrupción, o puede resultar una comunidad diferente. Las etapas más jóvenes de una serie (secuencia sucesional) son especialmente dinámicas en términos de cambio comunitario. Durante los primeros años de recuperación, la competencia no es intensa y predominan las especies ruderales seleccionadas por r. Las etapas posteriores de sucesión son mucho menos dinámicas y predominan las especies seleccionadas por K.
- Las microperturbaciones son perturbaciones locales que afectan a áreas pequeñas dentro de una comunidad intacta. Una microperturbación puede, por ejemplo, estar asociada con la muerte de un árbol grande individual, lo que resulta en una brecha en el dosel, por debajo de la cual el cambio comunitario es relativamente dinámico ya que las especies compiten para aprovechar la luz solar adicional. De manera similar, la muerte de una cabeza de coral individual representa una microperturbación dentro de una comunidad de arrecifes tropicales. Aunque los cambios ecológicos son dinámicos dentro de una brecha creada por una microperturbación reciente, a nivel de rodal la comunidad es relativamente estable. Las dinámicas comunitarias en fase de brecha ocurren en todos los ecosistemas, pero son especialmente importantes durante etapas posteriores de sucesión, como en los bosques de mayor crecimiento.

La variación espacial del entorno refleja el hecho de que las condiciones siempre están cambiando de un lugar a otro, y a veces extremadamente así. Estas variaciones espaciales influyen en el carácter de las comunidades ecológicas, de formas que pueden ser graduales o más rápidas:
- Los cambios graduales en las condiciones ambientales están asociados con la variación de la altitud en una montaña, diferencias de clima a grandes distancias entre continentes y otros gradientes relativamente continuos. Este tipo de cambio espacial se refleja en variaciones graduales de las comunidades debido a que las especies individuales tienen tolerancias y requisitos de condiciones ambientales diferentes pero superpuestos. Estas diferencias biológicas dan como resultado solapamientos de las distribuciones de especies, lo que puede dificultar que los ecologistas determinen las ubicaciones de los límites (ecotonos) entre tipos de comunidades.
- Los cambios rápidos en las condiciones ambientales ocurren en límites nítidos entre diferentes tipos de suelo o lecho rocoso, en las interfaces entre hábitats acuáticos y terrestres, y en lugares afectados por perturbaciones. Esta última influencia puede ocurrir, por ejemplo, entre un tracto forestal quemado y no quemado, o entre una reserva ecológica y su área circundante, que puede verse afectada por la agricultura o la silvicultura. Los cambios relativamente discretos en las condiciones ambientales favorecen grandes diferencias en los tipos de comunidad, con límites distintos entre ellos.
Ecología del Paisaje
Los paisajes (o paisajes marinos en el contexto marino) son un mosaico de “parches”, cada uno de los cuales representa una comunidad ecológica. Un paisaje puede contener diversos tipos de comunidades por las siguientes razones:
- cada comunidad refleja condiciones ambientales particulares, como diferentes tipos de suelo o lecho rocoso o variaciones de agua estancada (como en lagos, arroyos o humedales)
- las comunidades representan varias etapas en sucesión, como parches de diferente edad después de incendios forestales o daños por insectos
- las comunidades pueden estar relacionadas con el uso de la tierra, como cuando partes de los paisajes se ven afectadas por la urbanización, la agricultura, la silvicultura, las carreteras u otras influencias humanas.
Con el tiempo, los patrones espaciales de las comunidades en los paisajes son altamente dinámicos. Esto refleja en gran medida la influencia de las perturbaciones y la recuperación sucesional. Un parche que hoy es un pasto, un reciente corte claro o una quemadura puede ser un bosque maduro después de 50 años de sucesión. Del mismo modo, un estanque puede llenarse a lo largo de los siglos y convertirse en un humedal, que con más tiempo puede triunfar en un bosque. Los ecologistas utilizan el término “mosaico cambiante” para integrar las variaciones espaciales y temporales de las comunidades en los paisajes. Los siguientes factores afectan el mosaico cambiante de las comunidades.
- El tamaño del parche se relaciona con el área de rodales particulares de comunidades (un rodal es una comunidad en un lugar específico). Todas las especies necesitan un área mínima de hábitat para mantener sus poblaciones, y los pequeños parches pueden no ser adecuados para ese propósito. Sin embargo, parches relativamente pequeños pueden ayudar a mantener a una población que vive en varios rodales en el paisaje (una población extensa de este tipo se conoce como metapopulación). Esto puede suceder si los parches están conectados por corredores a otro hábitat adecuado, o si la especie es capaz de dispersarse a través del hábitat inhóspito circundante (para que esto ocurra, la matriz del paisaje debe ser permeable a los movimientos de la especie).
- La cantidad de borde es importante porque influye en la longitud del hábitat ecotono (transicional) asociado a un parche. Un parche circular tiene la menor relación de borde a área, y los parches más pequeños tienen relaciones más altas que los más grandes de la misma forma. Un ecotono entre tipos de parches es un tipo particular de hábitat, y puede ser utilizado selectivamente por “especies de borde”. Sin embargo, cuanto mayor es la relación de borde a área, menos hábitat “interior” hay (esto no está influenciado por las condiciones ecológicas asociadas a un ecotono). Los ecologistas han identificado “especies interiores” que tienen menos éxito si intentan usar hábitat cerca de un borde. Ciertas aves forestales, por ejemplo, experimentan mayores tasas de depredación y parasitismo en nidos en pequeños remanentes de bosque maduro (ver Capítulo 26).
- La conectividad se refiere a la presencia de vínculos entre parches discretos de hábitat similar. Estos eslabones pueden ser utilizados por una especie como corredores para moverse entre parches, permitiendo que su metapopulación funcione en el paisaje. Como se señaló anteriormente, la conectividad también está relacionada con la capacidad de una especie para dispersarse entre parches habitables a través del hábitat circundante.
- La adyacencia de clase de edad es importante en un paisaje en el que los parches representan diferentes etapas de una secuencia sucesional. Esto ocurre comúnmente en paisajes afectados por perturbaciones como incendios forestales, epidemias de insectos o tala clara. En general, los parches de una edad similar posterior a la perturbación serán comparables en muchos aspectos del hábitat, mientras que los de diferente edad serán menos similares. Esto puede ser una consideración importante para los movimientos de especies entre parches aislados que son adecuados como hábitat.
- Los requerimientos de hábitat complejos son característicos de algunos animales más grandes, como el venado, el oso y el lobo. Estas especies necesitan diferentes tipos de parches de hábitat para fines específicos en diversas épocas del año. Debido a que estos animales participan en diversas clases de comunidades, todos los parches de hábitat que necesitan deben estar presentes en el paisaje si se quiere sostener una metapopulación viable.
- La biodiversidad a nivel de paisaje está relacionada con la riqueza de los tipos de comunidad en un área extensa (ver Capítulo 7). Un paisaje cubierto uniformemente por una sola comunidad tiene menos biodiversidad a esta escala que uno compuesto por un mosaico rico y dinámico de diferentes tipos de comunidades.
- Las funciones a nivel de paisaje operan en áreas extensas y pueden integrar las influencias de muchos tipos de comunidades. Una cuenca hidrográfica, por ejemplo, es la extensión de terreno desde la cual el agua drena hacia un arroyo, lago o alguna otra masa de agua. La mayoría de las cuencas hidrográficas contienen diversos tipos de parches de hábitat, cada uno con influencias particulares en la hidrología y la química del agua. En general, las cuencas cubiertas de bosque maduro arrojan los flujos de agua más limpios. Otros servicios ambientales proporcionados por paisajes bien vegetados incluyen la evapotranspiración, el control de la erosión, la moderación de los extremos climáticos y la absorción de dióxido de carbono atmosférico y la liberación de oxígeno.
La ecología del paisaje es un área temática importante en la ciencia ambiental. Los humanos comúnmente afectan rodales individuales de determinados tipos de comunidades, pero muchos de los efectos ecológicos deben manejarse a escala de paisajes y paisajes marinos.

La Biosfera
La biosfera consiste en toda la vida y ecosistemas de la Tierra. Está delimitada por la presencia de organismos vivos, y es el único lugar en el universo que definitivamente se sabe que sustenta la vida. Los procesos ecológicos a nivel de la biosfera incluyen los regímenes climáticos, oceánicos y atmosféricos globales (Capítulo 3), el presupuesto energético planetario (Capítulo 4) y los ciclos globales de nutrientes (Capítulo 5). Estos procesos biosféricos influyen en toda la vida y los ecosistemas. Al mismo tiempo, la vida y los ecosistemas también influyen en los procesos a nivel de biosfera.
De hecho, algunos científicos han sugerido que puede haber cierto grado de control homeostático, o retroalimentación, entre las influencias recíprocas de los ecosistemas globales y su entorno. Una noción que describe estas relaciones biosfera-ambiente se conoce como la hipótesis de Gaia, una idea polémica popularizada por el científico James Lovelock. Sugiere que los organismos y ecosistemas han provocado que se produzcan grandes cambios en ciertos atributos físicos y químicos del medio ambiente global, y han dado como resultado mejoras en las condiciones de vida del planeta. La hipótesis contempla a todas las especies y ecosistemas de la Tierra como una especie de “superorganismo”, que se llama Gaia. Según Lovelock, Gaia intenta optimizar las condiciones ambientales para mejorar su propia salud y continuidad, y utiliza mecanismos de retroalimentación para ayudar a mantener condiciones dentro de un rango que la vida puede tolerar. Los antiguos griegos creían que Gaia (o Gaea) era el prolífico antepasado de muchos de sus dioses más importantes. Los romanos, que adoptaron muchos dioses e ideas griegas, conocían a Gaia como Terra. Más recientemente, el mito gaiano ha sido personificado como “madre Tierra”.
La idea gaiana es atractiva e interesante, en gran parte porque integra muchas ideas y observaciones a gran escala en una creencia consolidada y cosmovisión. No obstante, la Tierra es el único planeta del universo que se sabe que sustenta la vida y los ecosistemas, por lo que es la única réplica conocida en el gran experimento de la vida. En consecuencia, la hipótesis de Gaia no puede ser probada por experimentación rigurosa, y por esta razón muchos científicos rechazan sus inferencias. Excepto en los términos más amplios, las ideas gaianas pueden no ser útiles para ayudar a los humanos a manejar los impactos perjudiciales de su creciente población y actividades industriales en la biosfera. Sin embargo, algunas líneas intrigantes de evidencia pueden ser agrupadas en apoyo de la noción gaiana. Siguen dos ejemplos.
Oxígeno Atmosférico
La atmósfera primordial de la Tierra no contenía oxígeno (O 2). Este gas apareció sólo después de que evolucionaran los primeros organismos fotosintéticos, bacterias azul-verdes. Estas, y las algas verdes algo más tarde evolucionadas, desprenden el O 2 como un producto de desecho de la fotosíntesis. La concentración moderna de O 2 en la atmósfera, alrededor del 21%, ha resultado enteramente de la fotosíntesis y es un factor ambiental crítico para la mayoría de las especies y muchos procesos ecológicos clave. Parece que la concentración de O 2 atmosférico se ha mantenido bastante estable durante varios miles de millones de años. Esto sugiere un equilibrio a largo plazo entre la producción de O 2 por fotoautótrofos y su consumo por respiración, incluyendo la descomposición. Curiosamente, si la concentración de oxígeno fuera muy superior al 21%, digamos 25%, entonces la biomasa sería mucho más combustible. Esta condición conduciría a incendios forestales más frecuentes y extensos, lo que dañaría severamente los ecosistemas terrestres. Estas observaciones pueden interpretarse como que sugieren la existencia de un control homeostático de la concentración de O 2 atmosférico, operando a escala biosférica. Este control puede lograr un equilibrio entre la necesidad de tener suficiente O 2 para sostener los organismos más abundantes (que tienen un metabolismo aeróbico), y mayores concentraciones de O 2 que resultarían en conflagraciones destructivas.
El Efecto Invernadero
La concentración de dióxido de carbono (CO 2) en la atmósfera está regulada por un complejo de procesos físicos y biológicos por los cuales este gas es emitido y absorbido. El CO 2 atmosférico es importante en el efecto invernadero de la Tierra, que mantiene la temperatura superficial dentro de un rango que los organismos pueden tolerar (Capítulos 4 y 17). El efecto invernadero ayuda a mantener una temperatura superficial promedio de aproximadamente 15°C, en comparación con los -18°C que de otro modo se producirían y serían demasiado fríos para que los organismos la toleren. Los defensores de la hipótesis de Gaia sugieren que estas observaciones implican un control homeostático del CO 2 atmosférico y un control indirecto del efecto invernadero y el clima.
Existe evidencia clara de que los organismos y ecosistemas provocan cambios sustanciales en su entorno, y también que se ven afectados por esas condiciones. La comunidad científica, sin embargo, no apoya ampliamente la noción de que las especies y ecosistemas de la Tierra se han integrado de alguna manera en una simbiosis mutuamente benevolente dirigida a mantener una gama cómoda de condiciones ambientales.
La hipótesis de Gaia es, sin embargo, bastante útil en la ciencia ambiental. Las ideas gaianas enfatizan las diversas conexiones que existen dentro y entre los ecosistemas, así como las consecuencias dañinas de las acciones humanas que cada vez más están provocando grandes cambios ambientales y ecológicos. Si estos cambios excedieran los límites biosféricos de tolerancia y reparación homeostáticas, las consecuencias para la geofísiología y ecología del planeta podrían ser catastróficas.
Ecología Evolutiva
La ecología evolutiva es una fusión de ecología y evolución, implica la interpretación de las relaciones ecológicas en términos de evolución, selección natural y temas relacionados. Además, reconoce que las especies y su ecología tienen una historia evolutiva que implica cambios a lo largo del tiempo, generalmente ocurriendo como respuestas adaptativas a las influencias ambientales. La selección natural es una influencia particularmente significativa en el cambio evolutivo, y es evidente en los regímenes de factores estresantes y perturbaciones ambientales que afectan la supervivencia y el éxito reproductivo de los organismos. Las áreas temáticas centrales en la ecología evolutiva son la evolución de los rasgos de la historia de vida, de las relaciones entre especies (como mutualismo, parasitismo, polinización y depredador y presa), y de comunidades y biodiversidad en general. Ejemplos de la interpretación de la naturaleza en términos de ecología evolutiva incluyen convergencia y coevolución:
- La convergencia ocurre cuando especies no relacionadas ocupan nichos similares en ambientes distantes pero comparables. Como resultado son sometidos a regímenes paralelos de selección natural, y evolucionan para ser similares en morfología y comportamiento. Un ejemplo señalado anteriormente es el lobo placentario (Canis lupus) de Eurasia y Norteamérica y el lobo tilacino o marsupial (Thylacinus cynocephalus) de Tasmania.
- La coevolución ocurre cuando las especies interactúan de manera que afectan la supervivencia o el éxito reproductivo, de manera que están sujetas a regímenes de selección natural que resultan en un cambio evolutivo integrado. La intensidad de la coevolución puede, sin embargo, variar mucho. En casos extremos, puede conducir a la evolución de un mutualismo obligado, lo que significa que ninguno de los dos puede sobrevivir sin el otro. Los líquenes son un ejemplo, estos son mutualismos entre un hongo y una alga, ninguno de los cuales puede sobrevivir por sí solo.
La coevolución también afecta las relaciones de alimentación de los organismos. Por ejemplo, las plantas de algodoncillo (como el algodoncillo común, Asclepias syriaca) han evolucionado para producir altas concentraciones de glucósidos cardíacos, que son desagradables y venenosos para la mayoría de los animales y ayudan a proteger las plantas de ser consumidas. Sin embargo, la mariposa monarca (Danaus plexippus) ha evolucionado una tolerancia a esos bioquímicos y así puede comer tejidos de algodoncillo (de hecho, sus larvas no comen nada más). Cuando esto sucede, los glucósidos cardíacos se incorporan a los tejidos de los monarcas, dando una medida de protección contra ser comidos por aves y otros depredadores. La defensa es tan efectiva que la mariposa virrey no relacionada (Limenitis archippus) ha evolucionado un patrón de coloración y vuelo que imitan los del monarca, lo que ayuda a disuadir a sus propios depredadores.
También se han estudiado las interacciones coevolutivas entre muchas plantas y sus insectos polinizadores. Un ejemplo es la orquídea abeja (Ophrys apifera), cuya estructura floral ha evolucionado para parecerse a la forma del abdomen de la abeja solitaria Eucera. Esto induce a las abejas macho a tratar de copular con la orquídea, la cual se poliniza en el proceso (aunque también es capaz de autopolinizarse).

Algunos ecologistas incluso han especulado sobre la posibilidad de que la selección natural opere en niveles superiores a las poblaciones o especies (esto se conoce como selección de grupos). Por ejemplo, se ha especulado que ciertos ensamblajes boreales de bosques han evolucionado para ser altamente inflamables, y que esta característica ha promovido incendios forestales de reemplazo de pie, una perturbación que puede rejuvenecer el ecosistema. Es cierto que el bosque boreal maduro dominado por abeto negro o abedul blanco puede fácilmente encenderse y arder rápidamente, pero aún no existe una teoría que apoye la evolución adaptativa de las comunidades, a diferencia de sus especies constituyentes.
Como es cierto para toda la biología, la teoría evolutiva proporciona un elemento clave del trasfondo conceptual de la ecología. El conocimiento y las percepciones de la evolución y la ecología también son fundamentales para comprender los efectos ambientales de la economía humana.
Ecología Aplicada
La aplicación de principios ecológicos para tratar problemas económicos y ambientales se conoce como ecología aplicada (o como ecología ambiental). Hay tres áreas temáticas principales:
- el manejo de los recursos renovables, como los importantes en la agricultura, la pesca y la silvicultura
- la prevención o reparación de daños ecológicos, como los relacionados con la biodiversidad amenazada, y la restauración de tierras o aguas degradadas
- el manejo de procesos ecológicos, como la productividad, el almacenamiento de carbono, el ciclo de nutrientes, la hidrología y la erosión
Los capítulos posteriores de este libro de texto contienen muchos ejemplos del uso de la ecología aplicada para tratar problemas de manejo de recursos, contaminación y perturbación. Si bien el tema no se trata aquí en detalle, se puede ilustrar brevemente con los siguientes ejemplos:
- Establecer límites de cosecha: La tasa de productividad forestal en una región se puede estimar midiendo árboles en parcelas que se vuelven a muestrear a lo largo de los años, o sacando muestras de poblaciones de árboles y examinando el ancho de anillo anual de la madera recién cultivada. Esta información puede ser utilizada para establecer límites en la cantidad de madera que se puede cosechar sin degradar el recurso. Se pueden usar métodos análogos para medir la productividad para manejar la recolección sustentable de poblaciones de mamíferos, aves y peces.
- Aumento de la Productividad Biológica: La investigación ecológica aplicada puede determinar si se puede incrementar la productividad de los recursos biológicos, por ejemplo, si el crecimiento forestal podría mejorarse aplicando fertilizantes, aclarando rodales densos o estableciendo plantaciones. La investigación también puede predecir otras implicaciones de estas prácticas de manejo, incluidos los efectos sobre la biodiversidad y la calidad del agua.
- Remediación, Recuperación y Restauración: Los ecologistas pueden brindar asesoría basada en la investigación para mejorar las condiciones en áreas que han sido degradadas por contaminación o perturbación. Se pueden utilizar diversos tipos de esquemas para hacer frente a los daños ecológicos:
- La remediación implica acciones que se emprenden para enfrentar un problema particular, como encalar lagos y ríos para disminuir su acidez, plantar plantas tolerantes en ambientes contaminados, o emprender la cría y liberación en cautividad para aumentar la abundancia de una especie en peligro de extinción.
- La recuperación implica acciones más integrales para establecer un ecosistema productivo en tierras degradadas. Por ejemplo, un viejo relleno sanitario o un sitio industrial en desuso pueden ser recuperados a una cobertura permanente de vegetación, como un pasto.
- La restauración tiene el objetivo más elevado de intentar establecer un facsímil de automantenimiento de un ecosistema natural en tierras degradadas. Por ejemplo, las técnicas de restauración ecológica podrían ser utilizadas para convertir las tierras de cultivo abandonadas de nuevo en una pradera o bosque nativo.
- Compensación para Gases de Efecto Invernadero: Los ecologistas pueden predecir la cantidad de bosque que se debe cultivar y proteger para compensar las emisiones industriales de gases de efecto invernadero. Esto puede permitir a empresas y naciones reducir sus emisiones netas de CO 2 y avanzar hacia el cumplimiento de acuerdos internacionales para hacer frente al cambio climático.
- Reducción de la erosión: Algunas áreas han sido muy degradadas por la erosión causada por la deforestación y otros cambios en el uso del suelo. Los ecologistas pueden encontrar formas de lidiar con este problema, por ejemplo, plantando bosques, usando campos agrícolas en terrazas o administrando la hidrología local para reducir los flujos de agua terrestre o subsuperficial.
Conclusiones
La ecología es el estudio de las relaciones de los organismos con los factores ambientales que proporcionan los requisitos de vida y sustento. Estos factores incluyen recursos como nutrientes y alimentos, las influencias de otros organismos a través de la competencia y depredación, así como factores estresantes como perturbaciones y contaminación. El conocimiento de la ecología es fundamental para comprender muchos de los daños más importantes que la economía humana está causando a la biosfera. El conocimiento ecológico aplicado es esencial para gestionar los recursos renovables de manera sustentable, conservar la biodiversidad y evitar y reparar los daños causados por la contaminación y otras influencias destructivas de los humanos.
Preguntas para revisión
- Distinguir entre autecología y sinecología. Dar ejemplos para ilustrar cada uno.
- ¿Qué son la tasa de natalidad, tasa de mortalidad, tasa de inmigración y tasa de emigración? ¿Cómo influyen estos factores demográficos en los cambios en una población?
- ¿Qué influencias ambientales y biológicas afectan la estructura y función de una especie de comunidad ecológica con la que estás familiarizado?
- ¿Cuáles son los atributos de un paisaje ecológico (o paisaje marino)? Ilustra tu respuesta con un ejemplo en la región donde vives.
- En un sentido general, ¿cuáles son los objetivos y métodos de un proyecto en ecología de restauración?
Preguntas para Discusión
- Utilizar los principios de la autecología para discutir las necesidades de recursos y las tolerancias ambientales de los humanos.
- ¿Cómo se integra la economía humana a la biosfera?
- ¿Cómo son relevantes las ideas y los conocimientos sobre ecología poblacional para las personas?
- ¿Cuáles son los elementos centrales de la hipótesis de Gaia? ¿Qué evidencia existe para apoyar esta hipótesis? ¿Cómo son relevantes las ideas gaianas para la ciencia ambiental?
- Es difícil y costoso implementar un proyecto a gran escala en ecología de restauración. ¿Bajo qué tipo de condiciones crees que podría valer la pena un proyecto así?
Explorando problemas
- El nicho a veces ha sido descrito como la “ocupación” de una especie, lo que hace para ganarse la vida, los recursos que utiliza y su hábitat. Se le ha pedido que haga una presentación ante un grupo de no ecologistas, en la que se debe describir el nicho de los humanos. ¿Qué información incluirías en tu presentación?
Referencias citadas y lecturas adicionales
Begon, M., R.W. Howorth, y C.R. Townsend. 2014. Esenciales de la Ecología. 4ª ed. Wiley, Cambridge, Reino Unido.
Freedman, B. y J. Riley. 1980. Tendencias poblacionales de diversas especies de aves invernantes en el sur de Ontario. Biólogo de Campo Ontario, 34:49-79.
Freedman, B., J. Hutchings, D. Gwynne, J. Smol, R. Suffling, R. Turkington, R. Walker, y D. Bazeley. 2014. Ecología: Un contexto canadiense. 2a ed. Nelson Canadá, Toronto, ON.
Goodwin, C. E., B. Freedman, y S. M. McKay. 1977. Tendencias poblacionales en la invernada de aves acuáticas en la región de Toronto, 1929—1976. Biólogo de Campo Ontario, 31:1-28.
Grime, J.P. 2001. Estrategias de Plantas, Procesos Vegetales y Propiedades Ecosistémicas. 2a ed. J. Wiley, Londres.
Harris, J.A., P. Birch, y J. Palmer. 1996. Restauración y Recuperación de Tierras: Principios y Prácticas. Addison-Wesley, Lectura, MA.
Kimmins, J.P. 2003. Ecología Forestal. 3a ed. Prentice Hall, Nueva York, NY.
Krebs, C.J. 2008. Ecología: El análisis experimental de distribución y abundancia. 6a ed. Benjamin Cummings, Nueva York, NY.
Lovelock, J.E. 2000. Gaia: Una nueva mirada a la vida en la Tierra. Oxford University Press, Nueva York, NY.
MacDonald, G.M. y L.C. Cwynar. 1991. Tasas de crecimiento poblacional posglacial de Pinus contorta ssp. latifolia en el oeste de Canadá. Revista de Ecología, 79:417-429.
Mills, S. 1996. Al servicio de la naturaleza: Restaurando y rehabitando terrenos dañados. Beacon Press, Boston, MA.
Odum, E.P. 1997. Ecología: un puente entre la ciencia y la sociedad. Sinauer Associates, Nueva York, NY.
Odum, E.P. y G.W. Barrett. 2004. Fundamentos de Ecología. Brooks Cole, Florencia, KY.
Silverton, J. y D. Charlesworth. 2001. Introducción a la Biología de Poblaciones Vegetales. Blackwell Science Ltd, Oxford, Reino Unido.
Smith, L.E. 1991. Gaia: El crecimiento de una idea. St. Martin's, Nueva York.