
1.3: Dominios de la vida
- Page ID
- 54072

Creemos con razón que toda la vida en la tierra evolucionó a partir de una célula ancestral común que existió poco después de los orígenes de la vida en nuestro planeta. Hace demasiado tiempo, no toda la vida se dividía en dos grupos: ¡la verdadera bacteria y todo lo demás!
Ahora agrupamos la vida en uno de tres dominios:
Se encuentran entre los primeros descendientes de esa célula ancestral común. Carecen de núcleos (pro significado antes y karyon que significa núcleo, o núcleo). Incluyen bacterias y cianobacterias (algas verdeazuladas).
Incluir todas las formas de vida superiores, caracterizadas por células con núcleos verdaderos (Eu, verdadero; karyon, núcleo)
(que significa bacterias “viejas”) incluyen muchas bacterias extremófilas ('amantes' de la vida a temperaturas extremadamente altas, salinidad, etc.). Originalmente clasificadas como procariotas antiguas, se demostró que en 1990 las arquebacterias estaban separadas de procariotas y eucariotas, un tercer dominio de la vida.
Las arqueas se encuentran en ambientes tan inhóspitos como las aguas termales hirvientes o el hielo ártico, aunque algunas también viven en condiciones que son más templadas. Carl Woese comparó las secuencias de ADN de genes para ARN ribosómicos en bacterias normales y extremófilos. Con base en similitudes y diferencias de secuencia, concluyó que estas últimas son de hecho un dominio separado del resto de las bacterias así como de los eucariotas. Para una revisión, véase (Woese, C. 2004; Una nueva biología para un nuevo siglo. Microbiol. Mol. Biol. Rev. 68:173-186)
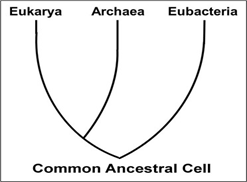
Los tres dominios de la vida (Archaea, Eubacteria y Eukarya) suplantaron rápidamente la división más antigua de los seres vivos en Cinco Reinos, la Monera (procariotas), Protista, Hongos, Plantas y Animales (¡todos eucariotas!) . En una sorpresa final, las secuencias de genes arquebacterianos indican claramente una ascendencia común de arqueas y eukarya.
¡Así, Archaea no son verdaderas bacterias! Comparten genes y proteínas así como vías metabólicas que se encuentran en eucariotas pero no en bacterias, apoyando su estrecha relación evolutiva con los eucariotas. Que también contienen genes y proteínas, así como vías metabólicas únicas para el grupo es un testimonio más de su estado de dominio. Comprender que todos los organismos vivos pertenecen a uno de los tres dominios ha cambiado drásticamente nuestra comprensión de la evolución. A continuación se ilustra la evolución de los tres dominios.
A. Los procariotas (Eubacteria = bacterias y C yanobacterias)
Las células procariotas carecen de núcleo y otros orgánulos como mitocondrias, cloroplastos, retículo endoplásmico y vesículas eucariotas variadas y membranas internas. Las bacterias sí contienen microcompartimentos bacterianos (BMC), pero estos están compuestos completamente de proteínas y no están rodeados por una membrana de fosfolípidos.
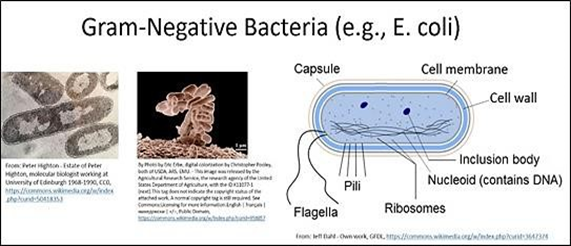
Estos funcionan por ejemplo en la fijación de CO2 para secuestrar metabolitos tóxicos para las células. Haga clic en Organelos Bacterianos para obtener más información. Las bacterias son típicamente unicelulares, aunque algunas (como algunas cianobacterias) viven vidas coloniales al menos en parte del tiempo. Las micrografías electrónicas de transmisión y barrido de bacterias en forma de varilla se muestran en el siguiente ejemplo a la izquierda. También se muestra un diagrama de la estructura bacteriana (derecha).
1. Reproducción Bacteriana
Sin los compartimentos que proporcionan los sistemas de membrana interna comunes a las células eucariotas, las químicas intracelulares, desde la replicación del ADN, la transcripción, la traducción y toda la bioquímica metabólica de la vida, ocurren en el citoplasma de la célula. El ADN es una doble hélice circular que se duplica a medida que la célula crece. Si bien no está encerrado en un núcleo, el ADN bacteriano se concentra en una región de la célula llamada nucleoide. Cuando no están abarrotadas a alta densidad, las bacterias replican su ADN a lo largo de la vida de la célula, dividiéndose por fisión binaria. El resultado es la partición igual de “cromosomas” bacterianos duplicados en nuevas células. El cromosoma bacteriano es esencialmente ADN desnudo, no asociado con proteínas.
2. Motilidad Celular y Posibilidad de un Citoesqueleto
El movimiento de las bacterias es típicamente por quimiotaxis, una respuesta a los químicos ambientales. Algunos pueden responder a otros estímulos como la luz (fototaxía). Pueden moverse hacia o lejos de nutrientes, sustancias nocivas/tóxicas, luz, etc., y lograr la motilidad de varias maneras. Por ejemplo, muchos se mueven usando flagelos compuestos en gran parte de la proteína flagelina. La flagelina está ausente en las células eucariotas. Por otro lado, el citoplasma de las células eucariotas está organizado por un complejo citoesqueleto de varillas y tubos hechos de proteínas actina y tubulina. Durante mucho tiempo se pensó que los procariotas carecían de estos o similares componentes citoesqueléticos. Sin embargo, recientemente se descubrieron dos genes bacterianos que codifican proteínas homólogas a la actina y tubulina eucariotas. La proteína MReB forma un anillo cortical en bacterias sometidas a fisión binaria, similar al anillo cortical de actina que pellizca las células eucariotas en división durante la citocinesis (la división real de una sola célula en dos células hijas más pequeñas). Esto se modela a continuación en la sección transversal (derecha) cerca de la mitad de una bacteria divisoria (izquierda).
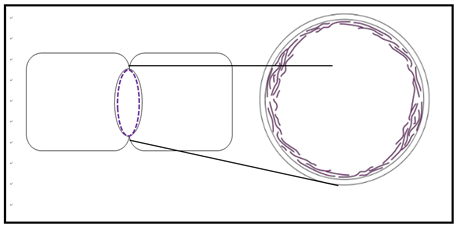
El gen ftsZ codifica un homólogo de proteínas tubulinas. Parece que junto con la flagelina, las proteínas MreB y FtsZ pueden formar parte de un citoesqueleto procariota primitivo involucrado en la estructura y motilidad celular.
3. Algunas bacterias tienen membranas internas
Mientras que las bacterias carecen de orgánulos (las estructuras unidas a la membrana de las células eucariotas), las membranas internas en algunas bacterias se forman como extensiones internas (invaginaciones) de la membrana plasmática. Algunos de estos capturan energía de la luz solar (fotosíntesis) o de moléculas inorgánicas (quimiolitotrofia). Los carboxisomas son vesículas fotosintéticas unidas a membrana en las que el CO2 se fija (reduce) en cianobacterias (se muestra a continuación).
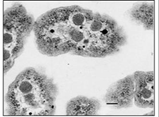
CC-BY; De: es.wikipedia.org/wiki/Archivo:Carboxisomes_em.jpg
Las bacterias fotosintéticas tienen sistemas de membrana interna menos elaborados.
4. Los ribosomas bacterianos hacen lo mismo que los ribosomas eucariotas... ¡y se parecen a ellos!
Los ribosomas son las máquinas sintetizadoras de proteínas de la vida. Los ribosomas de procariotas son más pequeños que los de los eucariotas, pero son capaces de traducir ARN mensajero eucariota (ARNm) in vitro. Subyacente a esta función básica común está el hecho de que los ARN ribosómicos de todas las especies comparten secuencias de bases y similitudes estructurales, lo que indica una larga relación evolutiva. Las similitudes de recuerdo revelaron la relación más estrecha de arqueas con eukarya que prokarya.
Claramente, las prokarya (Eubacterias) son un grupo diverso de organismos, ocupando casi todos los recovecos húmedos, secos, calientes o fríos de nuestro planeta. A pesar de esta diversidad, todas las células procariotas comparten muchas propiedades metabólicas estructurales y funcionales entre sí... ¡y con las arqueas y los eucariotas! Como hemos visto con los ribosomas, las propiedades estructurales y funcionales compartidas apoyan la ascendencia común de toda la vida. Por último, no solo compartimos ascendencia común con procariotas, incluso compartimos arreglos de vida con ellos. ¡Nuestras bacterias intestinales representan hasta 10 veces más células que las nuestras! Lee más en The NIH Human Microbiome Project. También consulta el siguiente enlace para Una relación entre microbiomas, dieta y enfermedad .
B. Las arquebacterias (Archaea)
Allessandro Volta, un físico que dio su nombre al Volt, ¡descubrió bacterias productoras de metano (metanógenos) en 1776! Los encontró viviendo en el ambiente extremo en el fondo del Lago Mayor, un lago compartido por Italia y Suiza. Estas bacterias inusuales son cheomoautótrofos que obtienen energía del H2 y CO2 y también generan gas metano en el proceso. No fue hasta la década de 1960 que Thomas Brock (de la Universidad de Wisconsin-Madison) descubrió bacterias termófilas que vivían a temperaturas cercanas a los 100oC en el Parque Nacional Yellowstone en Wyoming. Los organismos que viven en cualquier ambiente extremo pronto fueron apodados extremófilos. Una de las bacterias termófilas, ahora llamada Thermus aquaticus, se convirtió en la fuente de la polimerasa Taq, ¡la ADN polimerasa termoestable que convirtió a la reacción en cadena de la polimerasa (PCR) en un nombre familiar en laboratorios de todo el mundo!
Las bacterias extremófilas y “normales” son similares en tamaño y forma (s) y carecen de núcleos. Esto inicialmente sugirió que la mayoría de los extremófilos eran procariotas. Pero como demostró Carl Woese, ¡son las arqueas y la eukarya las que comparten una ascendencia común más reciente! Si bien algunas bacterias y eucariotas pueden vivir en ambientes extremos, las arqueas incluyen los extremófilos más diversos. Aquí hay algunos ejemplos de extremófilos:
• Acidófilos: crecen a pH ácido (bajo).
• Alcalífilos: crecen a pH alto.
• Halófilos: requieren altas concentraciones de sal; ver Halobacterium salinarium más abajo.

• Metanógenos: producen metano; arriba se muestra una sección transversal de Methanosarcina acetivorans (derecha). Obsérvese la ausencia de estructura interna significativa.
• Barófilos: crecen mejor a alta presión hidrostática.
• Psicrófilos: crecen mejor a una temperatura de 15 °C o inferior.
• Xerófilo s: crecimiento a muy baja actividad hídrica (condiciones de sequía o cerca de sequía).
• Termófilos e hipertermófilos: organismos que crecen mejor a 40°C o más, o 80°C o más, respectivamente. Pyrolobus fumarii, un hipertermófilo, puede vivir a una temperatura de 113°C; a continuación se muestra otro termófilo Thermus aquaticus, destacado por su papel en el desarrollo de la reacción en cadena de la polimerasa.

• Toxicolantes: crecen en presencia de altos niveles de elementos dañinos (e.g., charcas de benceno, desechos nucleares).
Las arqueas fueron originalmente vistas como rarezas de la vida, prosperando en entornos hostiles. También incluyen organismos que viven en ambientes menos extremos, incluyendo suelos, marismas e incluso en el colon humano. También son abundantes en los océanos donde son una parte importante del plancton, participando en los ciclos de carbono y nitrógeno. En las entrañas de vacas, humanos y otros mamíferos, los metanógenos facilitan la digestión, generando gas metano en el proceso. De hecho, ¡las vacas incluso han sido citadas como una de las principales causas del calentamiento global debido a sus prodigiosas emisiones de metano! En el lado positivo, las Archaea metanogénicas están siendo explotadas para crear biogás y tratar aguas residuales. Otros extremófilos son la fuente de enzimas que funcionan a altas temperaturas o en disolventes orgánicos. Como ya se señaló, algunas de ellas han pasado a formar parte de la caja de herramientas biotecnológicas.
C. Los eucariotas
1. Celdas grandes compartimentadas
El volumen de una célula eucariota típica es unas 1000 veces el de una célula bacteriana típica. La vida eucariota ni siquiera hubiera sido posible si no fuera por una división del trabajo de las células eucariotas entre diferentes orgánulos (estructuras unidas a la membrana). Imagina una bacteria como una habitación de 100 pies cuadrados (¡del tamaño de una habitación pequeña o un vestidor grande!) con una puerta. Ahora imagina una habitación 1000 veces más grande. Es decir, imagina una 'habitación' de 100,000 pies cuadrados. Se esperarían muchas habitaciones más pequeñas dentro de un espacio tan grande, cada una con su (s) puerta (s) propia (s). La célula eucariota se parece mucho a ese gran espacio, con muchas “habitaciones” interiores (es decir, orgánulos) con sus propias entradas y salidas. La “habitación” procariota más pequeña tiene una relación de área superficial/volumen de la membrana plasmática mucho mayor que una célula eucariota típica, lo que permite que los químicos ambientales requeridos entren y se difundan rápidamente por todo el citoplasma de la célula bacteriana. Por lo tanto, la comunicación química entre partes de una célula pequeña es rápida. Por el contrario, la comunicación sobre una mayor extensión de citoplasma dentro de una célula eucariota requiere las actividades coordinadas de los compartimentos subcelulares. Tal comunicación podría ser relativamente lenta. De hecho, las células eucariotas tienen menores tasas de metabolismo, crecimiento y reproducción que las células procariotas. La existencia de células grandes requirió la evolución de una división del trabajo apoyada en la compartimentación.
2. Descripción general de la estructura de células animales y vegetales
Las células animales y vegetales típicas con sus orgánulos y otras estructuras se ilustran a continuación.
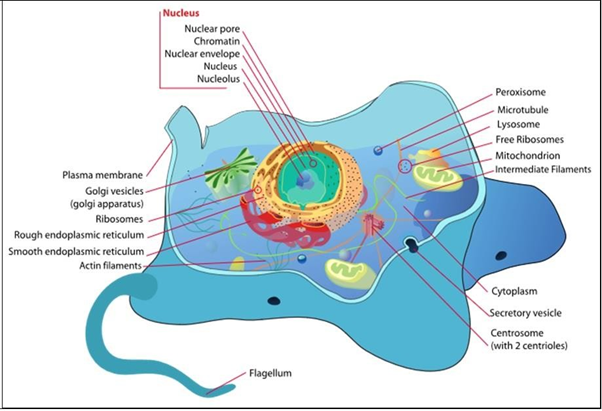
Una membrana plasmática (celular) rodea todas las células. Una pared celular rodea aún más células procariotas, algales, fúngicas y vegetales, creando una estructura rígida alrededor de la membrana celular y soportando la forma celular. Las paredes celulares bacterianas están compuestas por peptidoglicano, cadenas largas de polisacáridos unidas a cadenas polipeptídicas (aminoácidos). La celulosa, la hemicelulosa y la pectina son los principales polisacáridos de la pared celular de la planta. Las células fúngicas contienen una pared, cuyo componente principal es la quitina. La quitina es el mismo material que conforma el exoesqueleto o artrópodos (¡incluyendo insectos y langostas!). ¡Los hongos, más estrechamente relacionados con las células animales que con las plantas, son una bestia curiosa por varias razones! Por un lado, la organización de hongos y células fúngicas está algo menos definida que las células animales. Las estructuras entre células llamadas septos separan las hifas fúngicas, permiten el paso del citoplasma e incluso orgánulos entre las células. Algunos hongos primitivos tienen pocos o ningún septo, en efecto creando coenocitos, que son células gigantes únicas, con múltiples núcleos.
Terminamos esta mirada a los dominios de la vida señalando que, si bien los eucariotas son una minúscula minoría de todas las especies vivas, “su biomasa colectiva mundial se estima aproximadamente igual a la de los procariotas” (Wikipedia). Por otro lado, ¡nuestros cuerpos contienen 10 veces más células microbianas que células humanas! De hecho, cada vez es más claro que un humano debe tanto de su existencia a su microbiota (ver más arriba) como a sus células humanas. Teniendo en cuenta que las plantas y las células animales comparten muchas estructuras internas y orgánulos que realizan las mismas funciones o similares, veamos las mismas y describamos brevemente sus funciones.