14.4: Transposones desde McClintock
- Page ID
- 54276

Los transposones existen en todas partes donde miramos en procariotas y representan gran parte del ADN repetitivo eucariota. Como tales, pueden ser una gran proporción de genomas eucariotas, incluyendo algunos que ya ni siquiera se transponen. Los transposones alguna vez fueron considerados ADN inútil o basura, sin ninguna función obvia..., o genes egoístas sin otro propósito que la autorplicación. Pero a la luz de alguna nueva evidencia, ¡quizás no!
Como verá, los mecanismos de transposición comparten muchas características con la replicación, recombinación y reparación del ADN, e incluso la infección viral. A medida que estudias estos mecanismos, mantén en el fondo de tu mente que la transposición suele ser provocada por el estrés celular.
A. Secuencias de inserción (elementos IS)
Los elementos bacterianos IS fueron los primeros elementos móviles descritos después de los del maíz. Como veremos, comparten algunas características estructurales de los transposones eucariotas. Descubiertos a finales de la década de 1960, muchos han sido identificados (IS1, IS2..., IS10 etc.). Algunos se insertan en genes bien conocidos (por ejemplo, los del operón lac), pero la mayoría no lo son, probablemente porque hay poco ADN 'extra' en el genoma bacteriano compacto. Sin ADN extra no codificante para amortiguar las mutaciones dañinas, ¡pocas células bacterianas vivirían para contar una historia de transposición! Debería sorprendernos que los elementos IS puedan hacerse para transponerse en el laboratorio, pero generalmente son de naturaleza silenciosa.
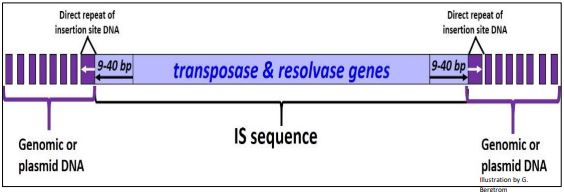
Los miembros de la familia de elementos IS varían en longitud de aproximadamente 750 a 1425 pb. Dentro de este tramo de ADN se encuentran genes transposasa y resolvasa cuyos productos son necesarios para la movilidad. En cada extremo del elemento IS se encuentran repeticiones invertidas, y cuando se encuentran en ADN genómico o plasmídico, la propia secuencia IS está flanqueada por repeticiones directas del genoma del huésped o ADN plasmídico que resultan del mecanismo de transposición. Nuevamente, debido a sus genomas compactos, las bacterias solo pueden tolerar un bajo número de copias de elementos IS en su genoma o en plásmidos (¡menos de diez copias y tan pocas como una!). A continuación se ilustra un elemento típico de IS.
B. Transposones compuestos: Elementos Tn
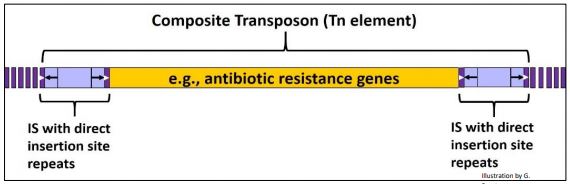
Si un par de elementos IS deben estar cerca uno del otro, separados por un tramo corto de ADN genómico o plasmídico, pueden transponerse juntos, transportando el ADN entre ellos como parte de un transposón compuesto, o elemento Tn. Si parte del ADN entre elementos IS en un elemento Tn contiene genes de resistencia a antibióticos, su transposición puede transportar y propagar estos genes a otro ADN en la célula. Los elementos Tn (como los elementos IS) están presentes en un número de copias bajo. A continuación se dibuja un elemento Tn genérico.
Los genes de resistencia a antibióticos preocupan a la comunidad médica; su propagación ha llevado a patógenos resistentes a los antibióticos que causan enfermedades que son cada vez más duras e incluso imposibles de tratar. Anteriormente vimos la 'transformación' genética de células estreptocócicas que recogen genes de virulencia en el ADN de células muertas. Transformamos rutinariamente células con plásmidos como parte de experimentos de ADN recombinante. Pero las bacterias pueden transferir ADN plasmídico entre ellas de manera bastante natural. Durante la conjugación bacteriana, un plásmido F (fertilidad) normalmente transfiere ADN entre tipos de apareamiento bacteriano compatibles (revise la conjugación bacteriana en otra parte de este texto para obtener más detalles). Un plásmido F que contiene un elemento Tn que alberga un gen de resistencia a antibióticos puede pasar de donante a receptor durante la conjugación. El elemento Tn puede transponerse al genoma bacteriano receptor. De esta manera, la transposición es una vía importante para la transferencia y propagación de la resistencia a los antibióticos.
C. Transposones complejos
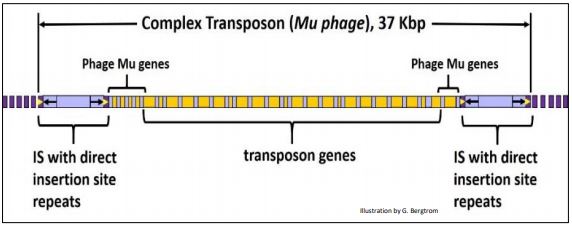
Los transposones complejos bacterianos también contienen otros genes además de los requeridos para la movilidad. Algunos transposones complejos se asemejan a un bacteriófago, o como en el caso del fago Mu, ¡en realidad son fagos! De hecho, Mu puede funcionar ya sea como un fago infeccioso que se reproduce en una célula infectada, o como un transposón en el genoma bacteriano. Los genes de transposón en el fago Mu se ilustran a continuación.
Después de infectar una bacteria, Mu puede entrar en la fase lítica de su ciclo de vida, replicando su ADN, produciendo y finalmente liberando nuevas 'partículas' de fagos infecciosos mediante la lisis de la célula bacteriana huésped. Alternativamente, al igual que otros fagos, Mu puede sufrir lisogenia, insertando su ADN en el cromosoma de la célula huésped. Las copias integradas de Mu podrían escindir y volver a entrar en la fase lítica para producir más fagos, particularmente si algún estrés ambiental amenaza la supervivencia bacteriana del huésped. Pero, una tercera opción de estilo de vida, la transposición, está disponible para Mu una vez que el fago se integra en el cromosoma bacteriano. Las tres opciones de estilo de vida para el fago Mu se ilustran en las siguientes páginas.
Las opciones de estilo de vida lítico y lisogénico para el fago Mu se muestran a continuación.
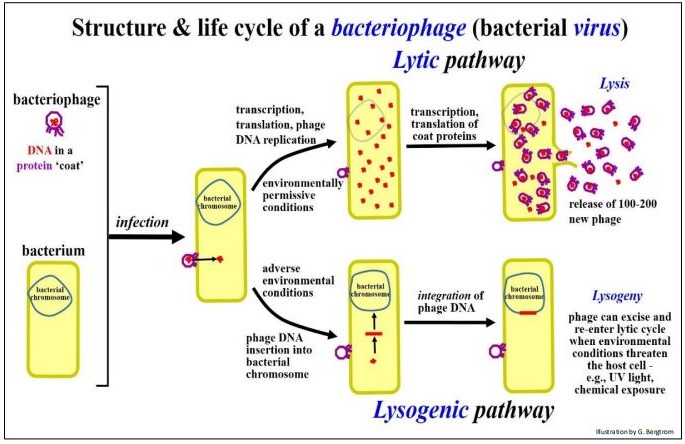
El ADN del fago mu puede actuar como un elemento transponible mientras se encuentra en la vía lisogénica, como se muestra a continuación
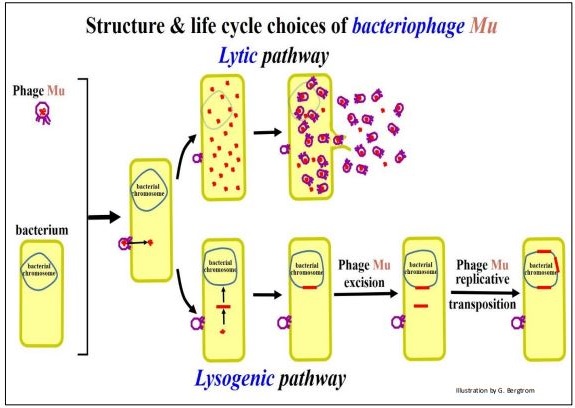
264 Elementos Móviles Bacterianos
Al pasar a una descripción de los transposones eucariotas, buscamos similitudes con los elementos bacterianos IS y Tn.
D. Visión general de los elementos transponibles eucariotas
Hay dos clases de transposones en eucariotas:
Clase I (Retrotransposones) se mover/'saltar' por transcripción de ARN en un locus, seguido de transcripción inversa e integración del ADNc de nuevo en ADN genómico en una ubicación diferente. Los retrotransposones pueden derivarse de (o ser la fuente de) retrovirus ya que los retrovirus activos se extraen e integran en el ADN de manera muy similar a los retrotransposones. Los retroposones son una subclase de retrotransposones (ver más abajo).
Clase II (Transposones de ADN) se mueven por uno de dos mecanismos. En la vía de cortar y pegar, el transposón deja un locus y se integra en otro. En la vía replicativa, el transposón original permanece en su lugar mientras las nuevas copias son móviles. La siguiente tabla muestra la distribución y proporción de genomas representados por diferentes clases/tipos de elementos transponibles.
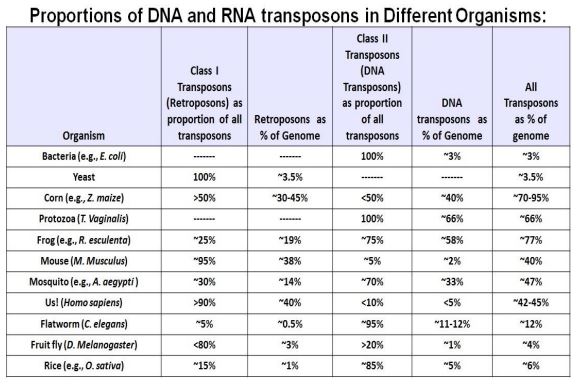
La tabla confirma que las bacterias contienen pocos transposones, mientras que los eucariotas varían ampliamente en la carga de transposones (transposones como porcentaje del ADN genómico), de tan baja como 4% a más de 70%.
En la siguiente tabla se resumen los elementos transponibles por clase, subtipo, tamaño, distribución genómica, mecanismo de transposición, etc.
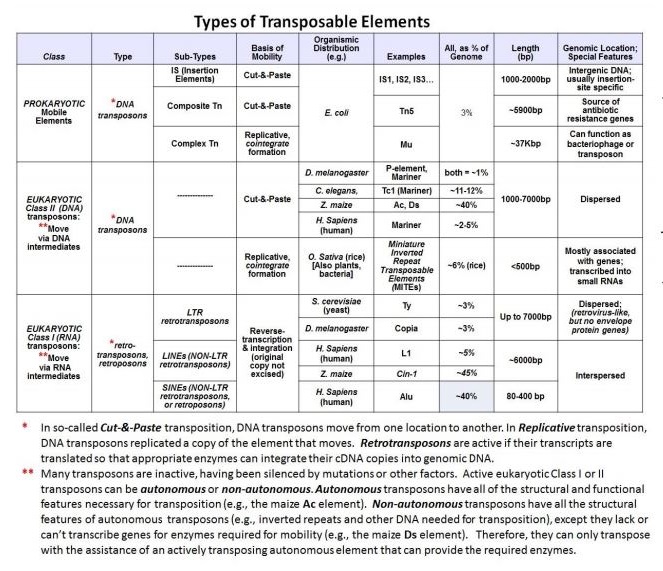
Entre las dos tablas anteriores, podemos concluir lo siguiente:
- La carga del transposón no se correlaciona con la complejidad evolutiva de los organismos.
- Los transposones compartidos tienen diferentes historias evolutivas en diferentes organismos.
- Donde los transposones permanecen activos, continúan dando forma a paisajes genómicos, especialmente en organismos con una alta carga de transposones.
Revisaremos algunas de estas conclusiones más adelante, después de analizar la estructura y el mecanismo de movilidad de los diferentes elementos transponibles.
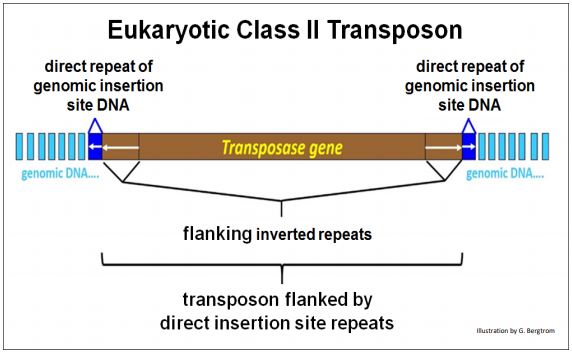
E. La estructura de los transposones de ADN eucariota (Clase II)
Los transposones de ADN eucariotas activos comparten características estructurales con elementos móviles bacterianos, incluidos los genes necesarios para la transposición, las repeticiones invertidas flanqueantes y las repeticiones directas flanqueantes del sitio de inserción del ADN de la célula hospedadora. A continuación se muestra la estructura característica de un transposón de ADN eucariota.
Los transposones de clase II pueden 'saltar' mediante mecanismos de corte y pegado o replicativos. La transposición de cortar y pegar elimina una copia de una ubicación y la mueve (transpone) a otra ubicación. Como su nombre indica, la transposición replicativa deja una copia del transposón original en su lugar mientras inserta una nueva copia en otra parte del genoma. La transposición mediante el mecanismo de cortar y pegar se muestra en un diagrama a continuación.
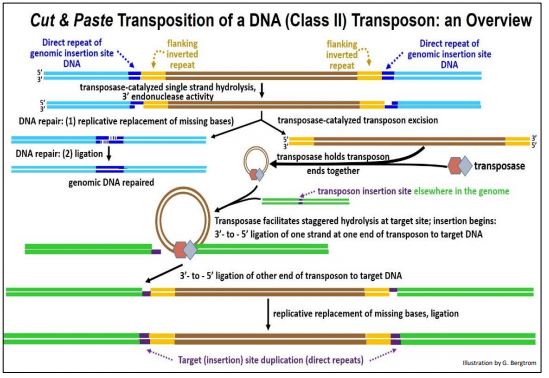
Tenga en cuenta que después de la transcripción del gen de la transposasa, la enzima mella el ADN y recorta los extremos 3'OH para crear un corte escalonado para extirpar el transposón. La transposasa realmente une los extremos del transposón durante la etapa de corte y media su inserción en un nuevo sitio de ADN. Después de la ligación de los extremos 3'OH del transposón al 5'OH en el sitio de inserción, la replicación reemplaza las bases faltantes, generando las repeticiones directas del ADN genómico de la célula huésped en el sitio de inserción. Un paso final de ligadura completa la transposición.
En la transposición replicativa, el transposón también mella y recorta el ADN en su sitio de inserción fuente (original). Pero, a diferencia del mecanismo de cortar y pegar, el transposón fuente no se extirpa.
A continuación se resumen los detalles del mecanismo replicativo de transposición.
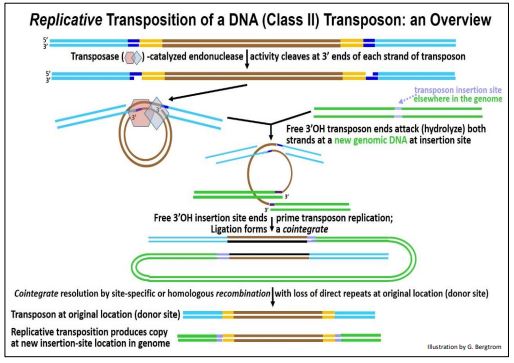
Después de cortar los extremos 3' del transposón en el sitio de inserción, la transposasa mantiene los extremos del transposón juntos mientras cataliza un ataque hidrolítico de ADN en un nuevo sitio de inserción. Esto es seguido por el cebado de la replicación de la cadena del transposón a partir de los extremos 3'OH de las cadenas de ADN del sitio de inserción. Se forma una estructura cointegrada en la que cada copia del transposón se ha realizado mediante replicación semi-conservadora. La cointegración se resuelve mediante uno de dos mecanismos de recombinación. El resultado deja copias del transposón tanto en el sitio original como en el nuevo sitio de inserción.
Comparemos y contrastemos las características de la transposición de ADN replicativo y de corte y pegado. Las características comunes son que:
- La transposasa codificada por transposón se une, une los extremos del transposón y cataliza la escisión monocatenaria (hidrólisis) dejando “extremos escalonados”.
- La transposasa mantiene los extremos del transposón juntos durante los pasos restantes.
Las diferencias entre los dos mecanismos son que en la transposición de corte y pasta, el transposón se extirpa completamente y luego se traspone a un nuevo sitio en el ADN genómico. Por el contrario, después de la escisión monocatenaria en transposición replicativa, los extremos 3' libres unidos a la transposasa del transposón hidrolizan ambas cadenas de ADN de cadena en un nuevo sitio de inserción. Después de la ligación de los extremos 3' de las cadenas del transposón a los extremos 5' de los extremos cortados del sitio de inserción del ADN genómico, los extremos 3' restantes de los extremos del ADN del sitio de inserción ceban la replicación del transposón, formando la cointegración, la cual es seguida por su resolución por una de las dos vías de recombinación.
F. Las estructuras de los transposones de ARN eucariota (Clase I)
Al igual que los transposones de ADN, todos los transposones de ARN dejan huellas en el sitio de inserción, es decir, repeticiones directas de ADN genómico que flanquean el elemento. A diferencia de los transposones de ADN, los transposones eucariotas activos de Clase I se mueven mediante un intermedio de ARN. También a diferencia de los transposones de ADN, carecen de repeticiones invertidas terminales.
La movilidad del ARN intermedio de todos los retrotransposones requiere un promotor que reconozca una enzima transcriptasa inversa así como enzimas endonucleasas e integrasas (que se describirán a continuación). Los transposones de ARN de Clase I autónomos incluyen retrotransposones LTR (por ejemplo, el elemento Ty de levadura) así como retrotransposones no LTR). Estas últimas incluyen las LÍNEAS autónomas (L ong I nterspersed N nuclear E lements). Las líneas autónomas LTR y no LTR contienen y expresan genes necesarios para las enzimas requeridas para la transposición. Por otro lado, los SINE (una subclase de retrotransposones no LTR) carecen de genes para las enzimas requeridas para la transposición y por lo tanto no pueden transponerse de manera independiente. Por lo tanto, son retrotransposones no autónomos que se basan en la actividad “verdadera” (autónoma) del retrotransposón para la movilidad. Los SINE a veces se llaman retroposones para distinguirlos de los retrotransposones autónomos.
249 Introducción a las características de la retrotransposición
A continuación, analizamos más de cerca las estructuras de Retrotransposón y los genes y actividades enzimáticas requeridos para la retrotransposición
1. Retrotransposones LTR: El Elemento Ty de Levadura
El transposón Ty alberga varios genes necesarios para la transposición. Estos incluyen:
- el gen Gag que codifica un antígeno específico de grupo, una proteína que forma una partícula similar a un virus que contendrá ADN de transposón transcrito inverso,
- el gen RT que codifica la transcriptasa inversa que hará copias transcritas inversamente de ARN transcritos retrotransposón.
- el gen Prt que codifica una proteasa que descompondrá la partícula similar a virus a medida que el retrotransposón ingresa al núcleo.
- el gen Int que codifica la integrasa requerida para la integración del retrotransposón en un sitio de inserción de ADN genómico.
A continuación se muestra un elemento Ty representativo ya que existiría integrado en el ADN genómico de levadura.
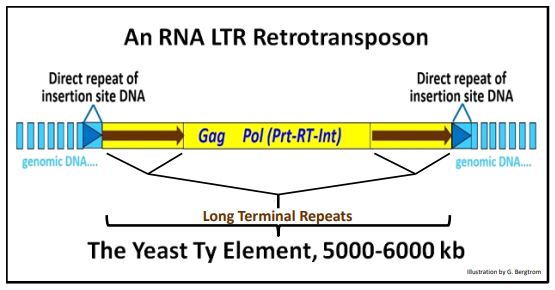
De hecho, muchos de los eventos en la transposición de Ty ocurren en la “partícula similar a virus” citoplásmica en células de levadura. Para ver más, da clic aquí. Nótese que la región Pol en la ilustración anterior consiste en marcos de lectura abiertos (ORF) superpuestos que codifican los genes Prt, RT e Int. El transposón listo para moverse consiste únicamente en la región del ADN simbolizada en amarillo.
2. Retrotransposones no LTR: LÍNEAS
LINES (L ong I nterspersed N nuclear E lements) también codifican enzimas necesarias para la transposición y, al igual que otros transposones, generan repeticiones directas del sitio diana flanqueando el elemento insertado. ¡Pero no tienen las largas repeticiones terminales! En cambio, sus ORF (genes) están flanqueados por regiones 5' y 3' no traducidas (UTR).
La estructura de la Línea L1 humana se dibuja a continuación.
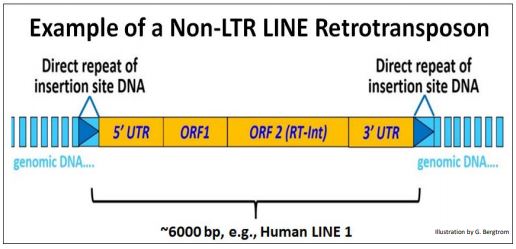
La UTR 5' contiene un promotor a partir del cual la ARN polimerasa II celular puede transcribir los genes aguas abajo (ver el capítulo Transcripción). El segundo de estos (ORF2) codifica la transcriptasa inversa y una actividad integrasa esencial para la transposición de la LINE. Todos los transposones autónomos Clase I (ARN-intermedio) comparten las siguientes características:
a) un Promotor en la 5' UTR a partir del cual puedan transcribirse.
b) una Transcriptasa Inversa que genera una copia de ADNc del elemento transponible.
c) RNasa H (una endonucleasa) que degrada ese transcrito después de la transcripción inversa.
d) Integrasa (como una transposasa) que cataliza la inserción de la copia del retrotransposón en los sitios de inserción.
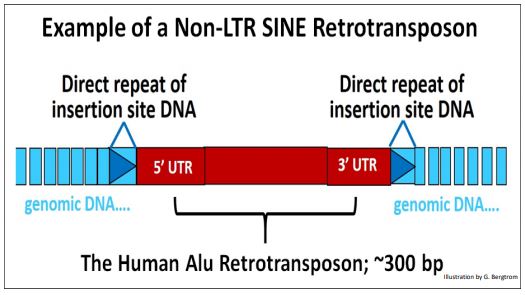
3. Retrotransposones SINE no LTR
Los retrotransposones SINE no LTR normalmente carecen de genes, pero su ADN no génico está flanqueado por UTR 5' y 3'. La ARN polimerasa III, que también transcribe los ARN de transferencia, también transcribe los SINE. Sin embargo, para transponer, se basan en la actividad concurrente de un transposón no LTR (un LINE) para proporcionar las actividades enzimáticas requeridas.
A continuación se muestra un SINE típico (por ejemplo, el elemento Alu).
G. Mecanismos de Retrotransposición
Existen dos mecanismos de retrotransposición: Retrotransposición Extracromosómicamente Primada (retrotransposones LTR por ejemplo) y Retrotransposición Imprimada en Sitio Objetivo de Inserción (Retrotransposones no LTR como LINES y SINEs). Estos serán considerados a continuación.
1. Retrotransposición Extracromosómicamente Imprimada (por ejemplo, de una LÍNEA)
Como su nombre indica, en retrotransposición extracromosómicamente cebada, una transcripción inversa circular del retrotransposón ataca, mella e integra en un sitio de inserción genómica. En este mecanismo, la transcriptasa inversa crea una copia de ADNc de un retro-elemento transcrito. La integrasa/endonucleasa luego se une a la copia de ADNc, manteniendo los extremos juntos, en efecto circularizándola. Esta ribonucleoproteína aislable se asemeja a un intasoma, una estructura similar al complejo de nucleoproteínas que cataliza la integración de ADNc retrovirales durante la lisogenia.
La retrotransposición extracromosómicamente imprimada se ilustra a continuación.
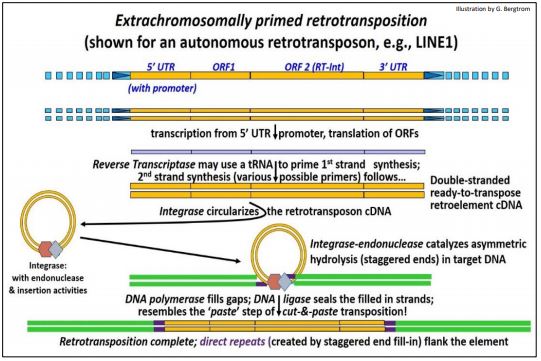
Recientemente se determinó la estructura tridimensional de un intasoma retroviral que interactúa con ADN y nucleosomas (para más información, ver Estructura 3D de Intasoma Retrovírico). De esta forma, el retrotransposón ataca el ADN en un sitio de inserción, creando extremos escalonados. Después de la inserción, se rellenan los huecos en el ADN. La ligadura sella el retrotransposón en su nueva ubicación, creando repeticiones directas del sitio de inserción.
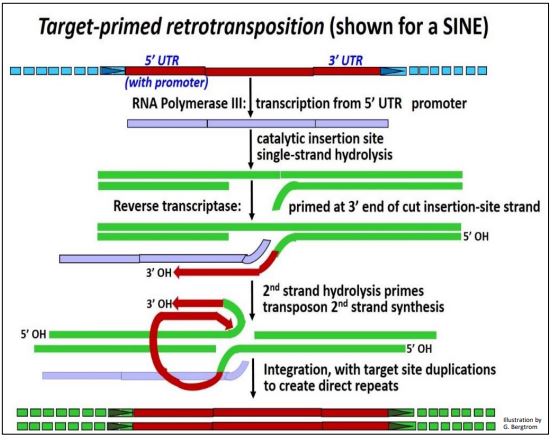
2. Retrotransposición SINE cebada en el sitio diana (por ejemplo, de un SINE)
Una característica clave de la retrotransposición cebada en el sitio diana (retroposición) es la ausencia de un transcrito inverso circular bicatenario unido a integrasa. En la transposición de SINE, la ARN polimerasa III (la misma enzima que cataliza la transcripción de ARNt y ARNr 5S) transcribe el SINE. Si se transcribe simultáneamente una LÍNEA, se elaborarán sus enzimas. Cuando su integrasa-endonucleasa cataliza la hidrólisis de una cadena de ADN en un nuevo sitio de inserción, el extremo 3'OH de esta cadena puede cebar la transcripción inversa de una cadena de ADNc de SINE por la transcriptasa inversa LINE. Después de la hidrólisis de la cadena de ADN del segundo sitio diana, su extremo 3'-OH ceba la replicación de la segunda cadena del ADNc de SINE. La integrasa completa la inserción de la Copy-sine en su nueva ubicación genómica. El mecanismo de retrotransposición cebado en el sitio diana de retrotransposición se ilustra a continuación.