
6.1: Fotosíntesis
- Page ID
- 53171

De una forma u otra, la energía de las moléculas de combustible de azúcar y grasa se deriva de la fotosíntesis -la conversión de la energía de la luz solar en energía de enlace químico, ya sea directamente en células vegetales fotosintéticas y ciertas bacterias fotosintéticas, o indirectamente por la ingestión de esas plantas y bacterias. La fotosíntesis es una idea simple: las moléculas de dióxido de carbono atmosférico se unen con moléculas de agua para formar azúcares y oxígeno:
\[\ce{CO2 + H2O ->[light] CH2O + O2}\]
La producción de energía utilizable a partir de la luz solar y la fijación del dióxido de carbono atmosférico son dos conjuntos separados de reacciones. En las plantas, la fotosíntesis se realiza únicamente en células que contienen cloroplastos. Los cloroplastos son orgánulos con un origen evolutivo que se sospecha que es similar al de las mitocondrias, y al igual que las mitocondrias, los cloroplastos generan ATP y utilizan un portador de electrones de alta energía basado en nicotinamidas. Hay más similitudes: ambas tienen membranas internas muy plegadas, aunque en los cloroplastos, hay tres membranas en total, mientras que las mitocondrias solo tienen dos. Finalmente, se incrusta una cadena de transporte de electrones en la membrana tilacoidea de los cloroplastos, funcionando de manera muy similar al transporte de electrones en las mitocondrias. Además de los componentes de transporte de electrones y ATP sintasa (estructural y funcionalmente casi idéntica a la ATP sintasa mitocondrial), la membrana tilacoidea también es rica en un conjunto de moléculas que no se encuentran en la membrana mitocondrial interna: moléculas pigmentarias absorbentes de luz.
En las plantas, estas moléculas de pigmento se dividen en dos clases: las clorofilas y los carotenoides (Figura\(\PageIndex{1}\)) pero solo las clorofilas pueden mediar en la fotosíntesis. Las bacterias fotosintéticas no contienen clorofila, pero sí tienen pigmentos carotenoides que pueden llevar a cabo la fotosíntesis. Ambos son hidrocarburos hidrofóbicos que se mantienen en su lugar dentro del plano de la membrana por proteínas transmembrana. Las clorofilas son fácilmente reconocibles por el anillo muy grande de porfirina que contiene Mg 2+, mientras que los carotenoides son cadenas largas de hidrocarburos que pueden tener o no pequeñas estructuras de anillo en los extremos (por ejemplo, β-caroteno). Si bien hay variación en la familia de la clorofila, todas ellas imparten un color verde a la hoja. Los carotenoides, por otro lado, tienen una gama mucho más amplia de colores desde amarillos hasta rojos. Tanto las clorofilas como los carotenoides son capaces de absorber energía luminosa de un rango de longitud de onda/energía particular y entrar en un estado excitado inestable. Cuando la molécula vuelve a su estado fundamental, la energía se emitiría como calor o luz en una situación aislada. Sin embargo, dentro del contexto de las matrices de pigmentos (complejo de antenas) en una célula viva, la mayor parte de la energía se traslada a otra molécula de pigmento de menor energía por transferencia de resonancia. Como se describe a continuación, solo un par de moléculas de clorofila en un complejo de antenas expulsará realmente un electrón a medida que cae de un estado excitado de nuevo al estado fundamental. Es la transferencia de ese electrón de alta energía el que alimenta la fotosíntesis.
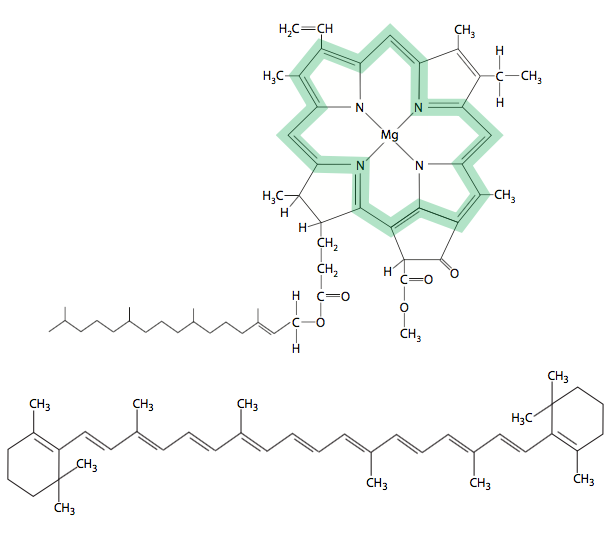
Las moléculas de clorofila están formadas por una cola hidrocarbonada de fitol que ancla la molécula dentro de una membrana, y un anillo de porfirina portador de electrones que contiene un catión de magnesio. Obsérvese que la cola de fitol no está dibujada a escala con el anillo de porfirina en la Figura\(\PageIndex{1}\). Entre los diferentes tipos de clorofila, los grupos químicos unidos al anillo pueden variar, y esta variación es responsable de diferencias en el espectro de absorción de un tipo de clorofila al siguiente. Por ejemplo, la clorofila a tiene picos de absorción a aproximadamente 430 y 662 nm, mientras que la clorofila b tiene picos a 453 y 642 nm. La diferencia entre los dos es pequeña: en C7, hay un grupo —CH 3 en la clorofila a, pero un grupo —CHO en la clorofila b. Actualmente, hay cinco clorofilas conocidas: la clorofila a se encuentra en todos los organismos fotosintéticos, la clorofila b solo se encuentra en plantas, las clorofilas c1 y c2 se encuentran en algas fotosintéticas, y clorofila d se encuentra en cianobacterias.
Los carotenoides tienen dos funciones. Como se señala en el texto primario a la izquierda, pueden participar en la transferencia de energía hacia las clorofilas del centro de reacción. También son una molécula protectora, previniendo la auto-oxidación del centro de reacción. Los carotenoides pueden ser eliminadores de radicales libres altamente eficientes debido a la conjugación de estructuras alternas de enlace de carbono simple y doble.
La fotosíntesis se puede dividir en dos mecanismos: las reacciones de luz, que utilizan la energía luminosa para excitar los electrones de ciertas clorofilas, y participan en la cadena de transporte de electrones para generar ATP y NADPH, y las reacciones oscuras, que utilizan ese ATP y NADPH para fijar el carbono de CO 2 a moléculas orgánicas (carbohidratos). Como su nombre lo indica, las reacciones lumínicas requieren energía lumínica para excitar a la clorofila y comenzar el transporte de electrones. Las reacciones oscuras, sin embargo, no requieren oscuridad. Son técnicamente independientes de la luz, pero en algunas plantas, las reacciones oscuras funcionan mejor a la luz por razones a discutir.
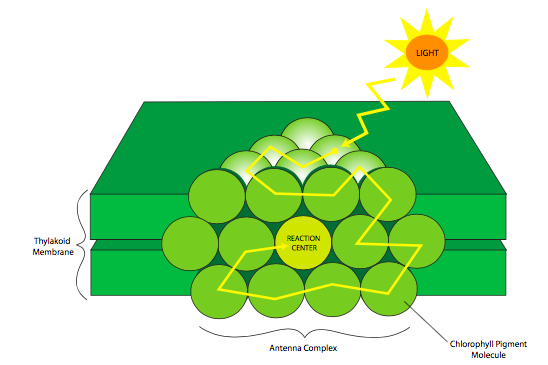
Las reacciones de luz están íntimamente ligadas a la anatomía de la membrana tilacoidea; específicamente, la disposición de las moléculas de pigmento que absorben la luz en los complejos de antena, también llamados complejos de recolección de luz (a veces abreviados LHC, que no deben confundirse con el Gran Colisionador de Hadrones). Estos pigmentos son retenidos por proteínas en grupos tridimensionales ordenados de manera que los pigmentos que absorben la luz de mayor energía están hacia la periferia, y las clorofilas de menor absorción de energía están en el centro (Figura\(\PageIndex{2}\)). La luz solar está compuesta por una amplia gama de longitudes de onda, algunas de las cuales son absorbidas transitoriamente por los pigmentos. Después de que una molécula de pigmento absorbe un fotón, la energía se libera y pasa a un pigmento sintonizado a un nivel de energía ligeramente inferior (longitud de onda más larga), y de ahí a un pigmento de menor energía, y así sucesivamente hasta que alcanza el centro de reacción clorofilas. De esta manera, la energía de una amplia gama de longitudes de onda/energías de luz puede contribuir a la producción de ATP y NADPH por las reacciones de luz. El complejo de antenas es crucial porque permite el uso de una mayor porción del espectro de luz solar. Y, como una matriz tridimensional apretada, los fotones que pasan por una molécula de pigmento bien pueden golpear a otra en su camino a través de la matriz. Todas estas características se combinan para aumentar la eficiencia del uso de la luz para la fotosíntesis. Las clorofilas del centro de reacción (P680 para el fotosistema II, P700 para el fotosistema I) son las únicas clorofilas que realmente envían electrones excitados a la cadena de transporte de electrones. Las otras clorofilas y pigmentos solo actúan para transferir la energía al centro de reacción.
Cuando se excita, el centro de reacción clorofila del fotosistema II (Figura\(\PageIndex{3}\)) inicia el proceso de transporte de electrones. Esta clorofila es parte de un complejo proteico que también incluye un complejo evolutivo de oxígeno basado en Mn (OEC), feofitina y un sitio de acoplamiento para plastoquinona. Si bien el electrón de clorofila es el excitado por la energía solar, el origen de los electrones para mantener la clorofila reabastecida proviene de la división (oxidación) del agua a O 2 y 4 H +.
El OEC, o complejo evolutivo de oxígeno (también WOC, complejo oxidante de agua) es una metaloenzima con un cúmulo catalítico Mn4oxCa, donde X es el número de m-oxo-puentes que conectan los átomos metálicos, con aminoácidos circundantes, especialmente tirosinas cruciales, desempeñando también un papel en la esfera de coordinación del activo sitio. El complejo general sufre una serie de 4 cambios de estado de oxidación ya que las clorofilas P680 son excitadas por la energía de la luz y los electrones de transferencia, pero en la actualidad no se sabe cuál es el estado exacto de oxidación de cualquier átomo de Mn dado a través de esta serie de cambios de estado. La reacción crucial es la formación del enlace O-O para formar O 2. Se proponen dos modelos para este mecanismo. Una es que el enlace O-O se forma cuando la OEC ha alcanzado su estado completamente oxidado, y un oxígeno en un estado radical m-oxo-puente interactúa con una molécula de agua. El otro mecanismo propuesto es que el enlace O-O se forma antes como un peróxido complejado mantenido por el centro OEC.
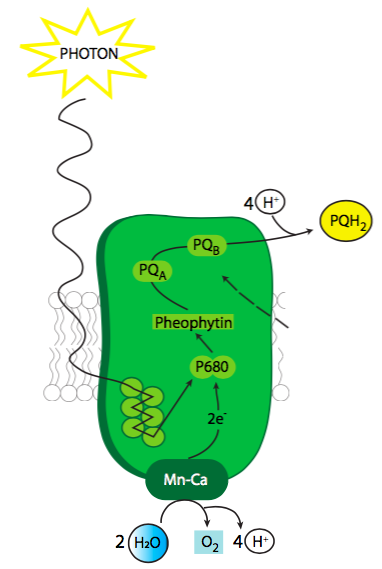
La cuestión de cómo una célula podría generar la energía necesaria para dividir el agua fue durante mucho tiempo un tema espinoso porque el agua es una molécula excepcionalmente estable. El modelo actual sugiere que la energía proviene de un oxidante extremadamente fuerte en forma de P680 +. Después de que P680 es energizado por la luz, un electrón excitado tiene suficiente energía para separarse de la clorofila y salta a feofitina. La feofitina se convierte en Pheo- temporalmente, y la separación de carga en el complejo entre P680 + y Pheo - ayuda a potenciar el poder oxidativo de P680 +. Esa atracción extraordinariamente fuerte para los electrones es lo que permite que la clorofila P680 los separe de H 2 O y divida el agua. De hecho, P680 + es uno de los oxidantes biológicos más fuertes conocidos. Dado que se deben tomar cuatro electrones para oxidar completamente dos moléculas de agua y generar oxígeno molecular, se necesitan cuatro eventos de fotoexcitación. Si bien el mecanismo exacto aún está por dilucidarse, parece que la OEC ayuda a estabilizar la molécula de agua durante este proceso así como a retener cada electrón a medida que sale.
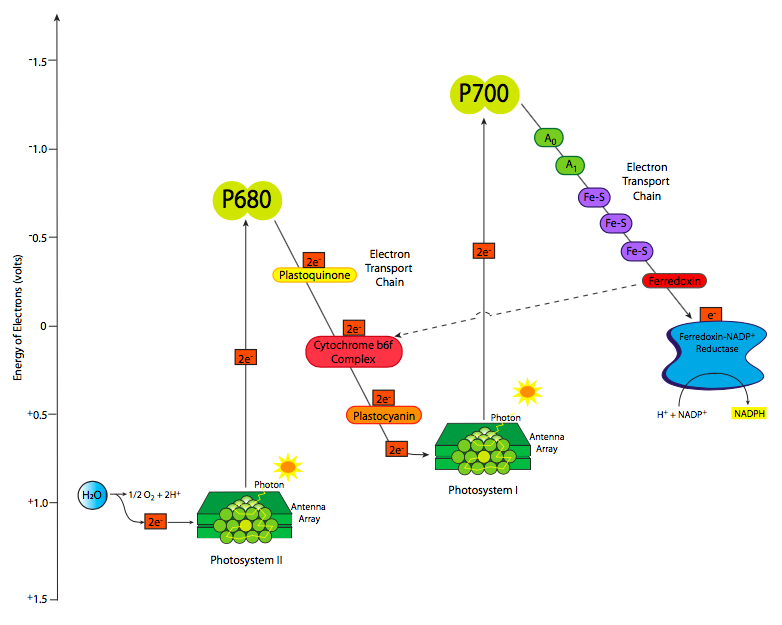
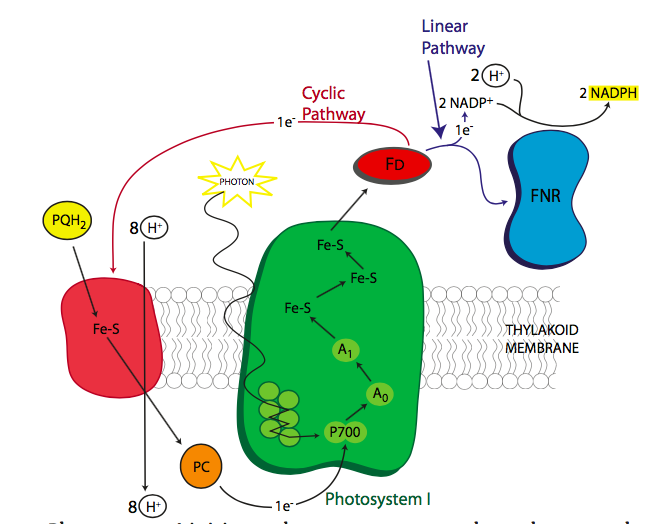
Los electrones excitados, pasando del OEC al P680 + a la feofitina, luego pasan al portador liposoluble, plastoquinona. La similitud del nombre con el portador mitocondrial ubiquinona no es una coincidencia. Funcionan de manera similar, y a medida que la plastoquinona toma los electrones, también toma protones del lado estromal de la membrana tilacoidea. El PQ se mueve dentro de la membrana de la feofitina al citocromo b 6 f. A medida que los electrones se transfieren al citocromo b 6 f, los protones se dejan caer en el lado luminal de la membrana, aumentando su concentración en el lumen del cloroplasto y construyendo un gradiente de protones a la potencia ATP sintasa. El citocromo b 6 f pasa los electrones a plastocianina, un portador de fase acuosa, que transporta los electrones a la clorofila del centro de reacción P700 del fotosistema I. Sin embargo, después de todas las transferencias, el nivel de energía de los electrones es ahora bastante bajo (Figura\(\PageIndex{4}\)) e incapaz de alimentar el próximas reacciones. Dado que ahora está en un centro de reacción clorofila, la respuesta obvia es volver a energizarla con un poco de luz solar. Esto eleva la energía electrónica lo suficiente como para reducir la ferredoxina. Ahora las cosas se complican un poco.
Esta parte de la fotosíntesis puede tomar una de dos direcciones, la vía lineal, que genera tanto NADPH como ATP, y la vía cíclica que genera principalmente ATP. La mayoría de las veces, se toma la vía lineal, con los electrones de la ferredoxina transferidos a través de Ferredoxina-NADPH reductasa (FNR) a NADPH. Sin embargo, a veces la célula requiere significativamente más ATP que NADPH, en cuyo caso, los electrones de la ferredoxina se transfieren de nuevo a plastoquinona vía ferredoxina-plastoquinona reductasa. Esto actúa tal como se describió anteriormente, y bombea más protones a través de la membrana para alimentar la ATP sintasa. La síntesis de ATP sube y la síntesis de NADPH baja.
El ATP y el NADPH generados por el cloroplasto son utilizados casi exclusivamente por el propio cloroplasto (y no distribuidos al resto de la célula) para alimentar las reacciones oscuras, que son energéticamente caras. De hecho, cuando las reacciones de luz no están funcionando debido a la oscuridad, algunas células vegetales tienen mecanismos para evitar que las reacciones oscuras utilicen los limitados recursos de respiración celular, no cloroplástica. El método más simple de dicha limitación es la sensibilidad al pH de rubisco (ribulosa bis-fosfato carboxilasa), al menos en plantas C3 (ver más abajo). Rubisco tiene un pH óptimo muy agudo a aproximadamente pH 8.0, por lo que mientras las reacciones de luz están funcionando y se bombean los protones, el pH sube a aproximadamente 8 y rubisco funciona, pero en la oscuridad, el pH vuelve a caer a su nivel basal cercano a 7.0, inhibiendo la actividad rubisco.