1.2: Introducción - Química Básica
- Page ID
- 53314

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
Fuente: BiochemFFA_1_2.pdf. Todo el libro de texto está disponible gratuitamente de los autores en http://biochem.science.oregonstate.edu/content/biochemistry-free-and-easy
“La química orgánica es la química de los compuestos de carbono. La bioquímica es la química de los compuestos de carbono que se arrastran” -Michael Adams.
Para entender la bioquímica, se debe poseer al menos una comprensión básica de la química orgánica y general. En esta breve sección, proporcionaremos una revisión rápida de los conceptos simples necesarios para entender la química celular. La química es química, ya sea en una célula o fuera de ella, pero la química biológica es un subconjunto particular de la química orgánica que a menudo involucra enormes macromoléculas, y eso ocurre en el ambiente acuoso de la célula.


Los enlaces covalentes, como ustedes saben, son el resultado del reparto de electrones entre dos átomos. Los enlaces iónicos, por el contrario, se forman cuando un átomo dona un electrón a otro, como en la formación de cloruro de sodio. Los enlaces covalentes simples pueden rotar libremente, pero los dobles enlaces no pueden. Los enlaces simples alrededor de un átomo de carbono están dispuestos en un tetraedro con ángulos de enlace de 109.5° entre sí, con el carbono en el centro (Figura 1.19). Los carbonos de doble enlace crean una estructura plana con ángulos de unión típicamente de aproximadamente 120°.
Electronegatividad
La electronegatividad es una medida de la afinidad que tiene un núcleo por los electrones de la capa externa (Cuadro 1.2). La alta electronegatividad corresponde a una alta afinidad. Los electrones en un enlace covalente se mantienen más cerca del núcleo con una mayor electronegatividad en comparación con un núcleo con menor electronegatividad.
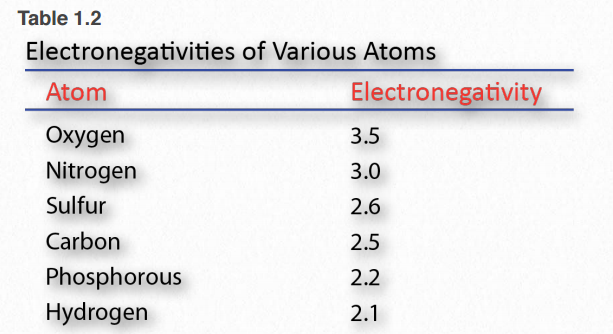
Cuadro 1.2 Imagen de Aleia Kim
Por ejemplo, en una molécula de agua, con hidrógeno unido covalentemente al oxígeno, los electrones son “arrastrados” hacia el oxígeno, que es más electronegativo. Debido a esto, hay una carga negativa ligeramente mayor cerca del átomo de oxígeno del agua, en comparación con el hidrógeno (que, correspondientemente tiene una carga positiva ligeramente mayor). Esta distribución desigual de carga configura un dipolo, siendo un lado algo negativo y el otro algo positivo. Debido a esto, la molécula se describe como polar.
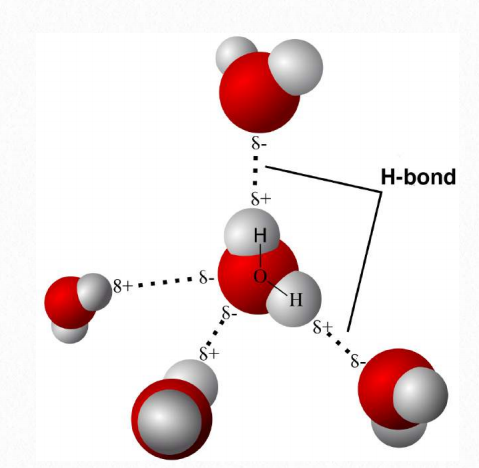
Los enlaces de hidrógeno entre las moléculas de agua son el resultado de la atracción de las cargas parciales positivas y negativas parciales sobre diferentes moléculas de agua (Figura 1.20). También se pueden formar enlaces de hidrógeno entre los hidrógenos con una carga positiva parcial y otros átomos fuertemente electronegativos, como el nitrógeno, con una carga parcial negativa. Es importante recordar que los enlaces de hidrógeno son interacciones entre moléculas (o partes de moléculas) y no son enlaces entre átomos, como enlaces covalentes o iónicos. Los enlaces entre hidrógeno y carbono no forman cargas parciales significativas debido a que las electronegatividades de los dos átomos son similares. En consecuencia, las moléculas que contienen muchos enlaces carbono-hidrógeno no formarán enlaces de hidrógeno y por lo tanto, no se mezclan bien con el agua. Tales moléculas se llaman hidrofóbicas. Otros compuestos con la capacidad de hacer enlaces de hidrógeno son polares y pueden disolverse en agua. Se les llama hidrófilos. Las moléculas que poseen ambas características se denominan anfifílicas.
Interacciones débiles
Los enlaces de hidrógeno son un tipo de interacción electrostática (es decir, basada en la carga) entre dipolos. Otras formas de interacciones electrostáticas que son importantes en la bioquímica incluyen interacciones débiles entre una molécula polar y un dipolo transitorio, o entre dos dipolos temporales. Estos dipolos temporales resultan del movimiento de electrones en una molécula. A medida que los electrones se mueven, el lugar donde se encuentran, en un momento dado, se vuelve temporalmente más cargado negativamente y ahora podría atraer una carga positiva temporal sobre otra molécula. Como los electrones no se quedan quietos, estos dipolos son de muy corta duración. Así, la atracción que depende de estos dipolos fluctúa y es muy débil. Las interacciones débiles como estas a veces se llaman fuerzas de van der Waals. Muchas interacciones moleculares en las células dependen de interacciones débiles. Aunque los enlaces de hidrógeno individuales u otras interacciones dipolo-dipolo son débiles, debido a su gran número, pueden resultar en interacciones bastante fuertes entre moléculas.
Oxidación/reducción
La oxidación implica pérdida de electrones y la reducción da como resultado ganancia de electrones. Por cada oxidación biológica, hay una reducción correspondiente: una molécula pierde electrones por otra molécula. Las reacciones de oxidación tienden a liberar energía y son una fuente de bioenergía para las células quimiotróficas.
Ionización
La ionización de biomoléculas, por el contrario, no implica oxidación/reducción. En la ionización, un ion hidrógeno (H+) deja atrás su electrón a medida que sale (dejando atrás una carga negativa) o se une a un grupo (agregando una carga positiva). Las ionizaciones biológicas implican típicamente grupos carboxilo o aminas, aunque también pueden ionizarse fosfatos o sulfatos. Un grupo carboxilo puede tener dos estados de ionización: una carga de -1 corresponde al carboxilo sin su protón y una carga de cero corresponde a la carga del carboxilo con su protón encendido. Una amina también tiene dos estados de ionización. Una carga de cero corresponde a un nitrógeno con tres enlaces covalentes (generalmente en forma de C-NH2) y una carga de +1 corresponde a un nitrógeno que hace cuatro enlaces covalentes (generalmente X-NH3 +).
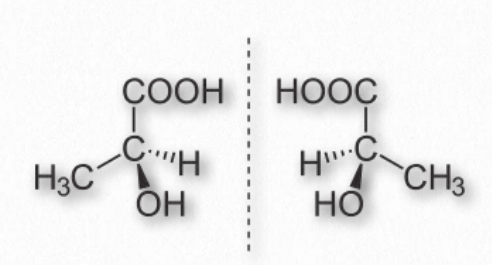
Estereoquímica
Un carbono tiene la capacidad de hacer cuatro enlaces simples (formando una estructura tetraédrica) y si se une a cuatro grupos químicos diferentes, sus átomos pueden disponerse alrededor del carbono de dos maneras diferentes, dando lugar a una “mano” estereoquímica (Figura 1.21). Cada carbono con tal propiedad se conoce como un centro asimétrico. La propiedad de la mano solo ocurre cuando un carbono tiene cuatro grupos diferentes unidos a él. Las enzimas tienen estructuras 3-D muy específicas, por lo que para las moléculas biológicas que pueden existir en diferentes formas estereoisoméricas, una enzima que la sintetiza produciría solo uno de los posibles isómeros. Por el contrario, las mismas moléculas elaboradas químicamente (no usando enzimas) terminan con cantidades iguales de ambos isómeros, llamadas mezcla racémica.
Energía libre de Gibbs
El cálculo de energía libre de Gibbs nos permite determinar si una reacción será espontánea, tomando en consideración dos factores, el cambio en la entalpía (ΔH) y el cambio en la entropía (ΔS). El contenido de energía libre de un sistema viene dado por la energía libre de Gibbs (\(G\)) y es igual a la entalpía (\(H\)) para un proceso menos la temperatura absoluta (T) por la entropía (S)
\[G = H = TS\]
Para un proceso, el cambio en la energía libre de Gibbs ΔG viene dado por
\[ΔG = ΔH - TΔS\]
Un negativo\(ΔG\) corresponde a la liberación de energía libre. Las reacciones que liberan energía son exergónicas, mientras que las que absorben energía se llaman endergónicas.
El cambio de energía libre de Gibbs estándar biológico (ΔG°') corresponde al ΔG para un proceso bajo condiciones estándar de temperatura, presión y pH = 7. Para una reacción
\[aA + bB \rightleftharpoons cC + dD,\]
la constante de equilibrio,\(K_{eq}\) es igual a
\[K_{eq} = \dfrac{ [C]^c_{eq} [D]^d_{eq}}{[A]^a_{eq} [B]^b_{eq}}\]
donde\(a\),\(b\),\(c\), y\(d\) son enteros en la ecuación balanceada. Grandes valores de\(K_{eq}\) corresponden a reacciones favorables (más C y D producidas que A y B) y pequeños valores de\(K_{eq}\) media lo contrario. En equilibrio,
\[ΔG^{o\prime} = -RT \ln K_{eq}\]
Si un proceso tiene a\(ΔG = Z\) y un segundo proceso tiene a\(ΔG = Y\), entonces si los dos procesos están vinculados,\(ΔG\) y\(ΔG^{o \prime} \) los valores para la reacción global serán la suma de los valores individuales ΔG y ΔG°'.
\[ΔG_{total} = ΔG_1+ ΔG_2 = Z + Y\]
\[ΔG^{o \prime }_{total} = ΔG_1^{o \prime}+ ΔG_2^{o\prime}\]
Catálisis
La catálisis es un aumento en la velocidad de una reacción inducida por una sustancia que, en sí misma, no cambia por la reacción. Debido a que los catalizadores permanecen inalterados al final de una reacción, una sola molécula de catalizador se puede reutilizar durante muchos ciclos de reacción. Las proteínas que catalizan reacciones en las células se denominan enzimas, mientras que las ribozimas son moléculas de ARN que actúan como catalizadores.