
5.3: Energía - Fotofosforilación
- Page ID
- 53046

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Fuente: BiochemFFA_5_3.pdf. Todo el libro de texto está disponible de forma gratuita de los autores en http://biochem.science.oregonstate.edu/content/biochemistry-free-and-easy

Fotofosforilación
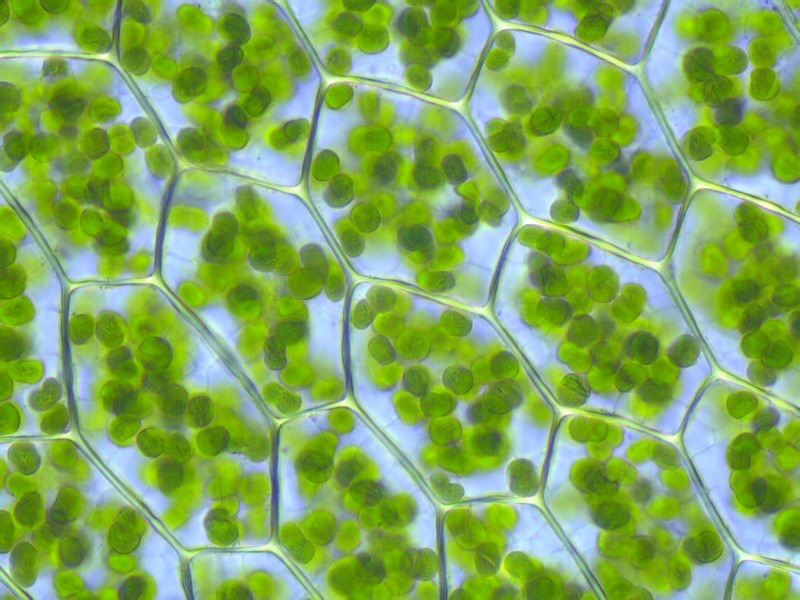
El tercer tipo de fosforilación para producir ATP se encuentra únicamente en las células que realizan la fotosíntesis. Este proceso es similar a la fosforilación oxidativa de varias maneras. Una diferencia primaria es la última fuente de energía para la síntesis de ATP. En la fosforilación oxidativa, la energía proviene de electrones producidos por la oxidación de moléculas biológicas. En la fotosíntesis, la energía proviene de la luz del sol. Los fotones del sol interactúan con moléculas de clorofila en centros de reacción en los cloroplastos (Figuras\(\PageIndex{1}\) y\(\PageIndex{2}\)) de plantas o membranas de bacterias fotosintéticas.
Las similitudes de la fotofosforilación con la fosforilación oxidativa incluyen:
- una cadena de transporte de electrones asociada a membrana
- creación de un gradiente de protones
- cosechar energía del gradiente de protones haciendo ATP con la ayuda de una ATP sintasa.
Algunas de las diferencias incluyen:
- la fuente de los electrones — H2O para la fotosíntesis versus NADH/FADH2 para la fosforilación oxidativa
- dirección del bombeo de protones: hacia el espacio tilacoideo de los cloroplastos versus fuera de la matriz de la mitocondria
- movimiento de protones durante la síntesis de ATP — fuera del espacio tilacoide en la fotosíntesis versus hacia la matriz mitocondrial en la fosforilación oxidativa
- naturaleza del aceptor de electrones terminal — NADP+ en fotosíntesis versus O2 en fosforilación oxidativa.
Transporte de electrones: cloroplastos vs mitocondrias
De alguna manera, el movimiento de electrones en los cloroplastos durante la fotosíntesis es opuesto al del transporte de electrones en las mitocondrias. En la fotosíntesis, el agua es la fuente de electrones y su destino final es NADP+ para hacer NADPH. En las mitocondrias, NADH/FADH2 son fuentes de electrones y H2O es su destino final. ¿Cómo hacen los sistemas biológicos que los electrones vayan en ambos sentidos? Parecería ser el equivalente a ir y venir de un lugar en particular mientras siempre se va cuesta abajo, ya que los electrones se moverán según el potencial.
Energía solar
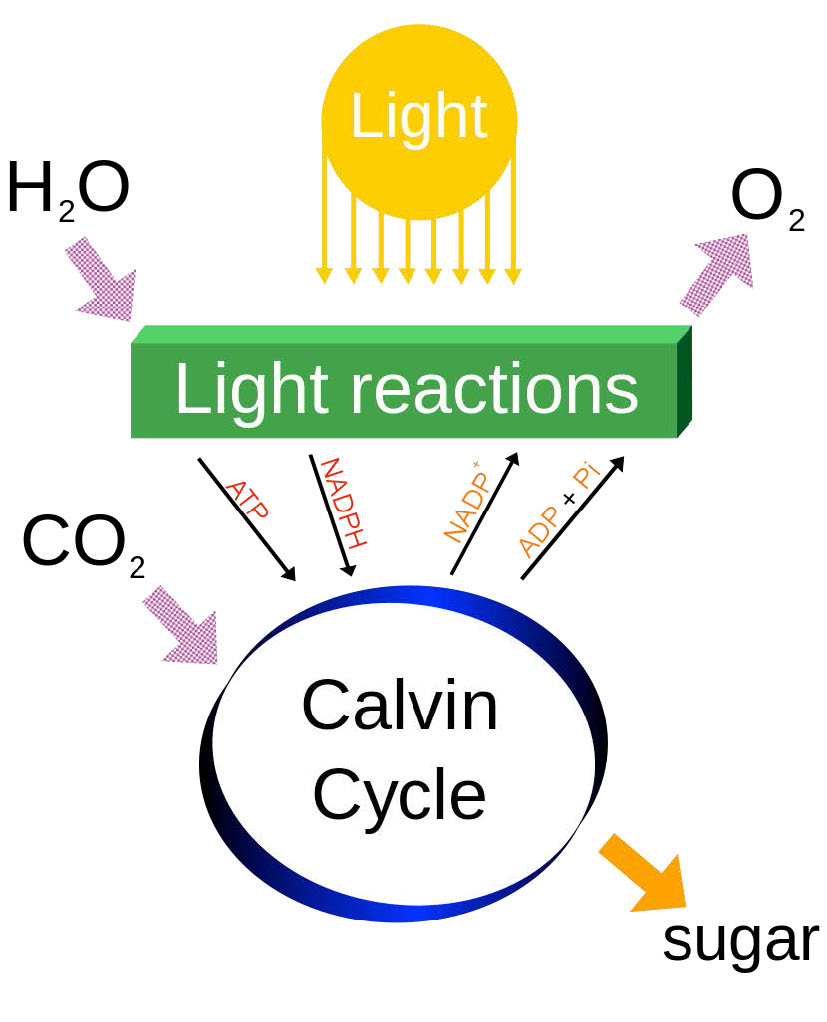
La respuesta es la energía capturada de los fotones del sol (Figura 5.59), que eleva los electrones a una energía donde se mueven “cuesta abajo” hacia su destino NADPH en un esquema en forma de Z. El movimiento de electrones a través de este esquema en las plantas requiere energía de fotones en dos lugares para “levantar” suficientemente la energía de los electrones.
Por último, cabe señalar que la fotosíntesis en realidad tiene dos fases, referidas como el ciclo de luz (descrito anteriormente) y el ciclo oscuro, que es un conjunto de reacciones químicas que captura CO2 de la atmósfera y lo “fija”, en última instancia, en glucosa. El ciclo oscuro también se conoce como el Ciclo Calvino y se discute AQUÍ.
Fotosíntesis
La fotosíntesis es un proceso de captura de energía que se encuentra en plantas y otros organismos para cosechar energía lumínica y convertirla en energía química. Esta energía fotoquímica se almacena en última instancia en carbohidratos que se elaboran con ATP (de la recolección de energía), dióxido de carbono y agua. En la mayoría de los casos, un subproducto del proceso es el oxígeno, que se libera del agua en el proceso de captura. La fotosíntesis es responsable de la mayor parte del oxígeno en la atmósfera y suministra los materiales orgánicos y la mayor parte de la energía utilizada por la vida en la Tierra.
Pasos
Los pasos en el proceso de fotosíntesis varían ligeramente entre organismos. En una visión amplia, siempre comienza con la captura de energía de la luz por complejos proteicos, que contienen pigmentos de clorofila, llamados centros de reacción. Las plantas secuestran estas proteínas en cloroplastos, pero las bacterias, que no tienen orgánulos, las incrustan en sus membranas plasmáticas.
La energía de la luz se utiliza para separar electrones de los donantes de electrones (generalmente agua) y dejar un subproducto (oxígeno, si se utilizó agua). Los electrones son donados a un portador y finalmente son aceptados por el NADP+, para convertirse en NADPH. A medida que los electrones viajan hacia el NADP+, generan un gradiente de protones a través de la membrana tilacoidea, que se utiliza para impulsar la síntesis de ATP. Así, el NADPH, el ATP y el oxígeno son los productos de la primera fase de la fotosíntesis llamadas reacciones de luz. La energía del ATP y los electrones del NADPH se utilizan para reducir el CO2 y construir azúcares, que son el último almacenamiento de energía que surge directamente de la fotosíntesis.
Cloroplastos
Los cloroplastos se encuentran en casi todas las células vegetales sobre el suelo, pero se concentran principalmente en las hojas. El interior de una hoja, debajo de la epidermis, está conformado por tejido de fotosíntesis llamado mesófilo, que puede contener hasta 800,000 cloroplastos por milímetro cuadrado.
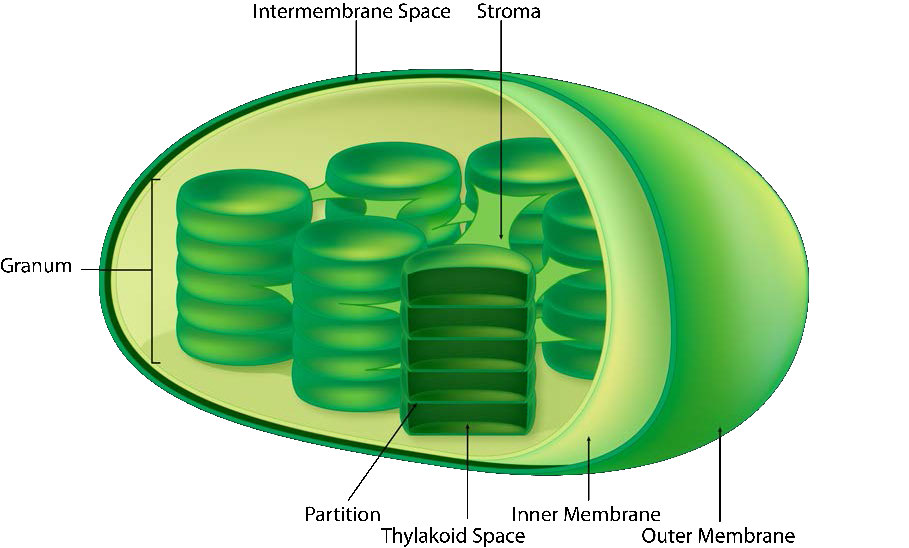
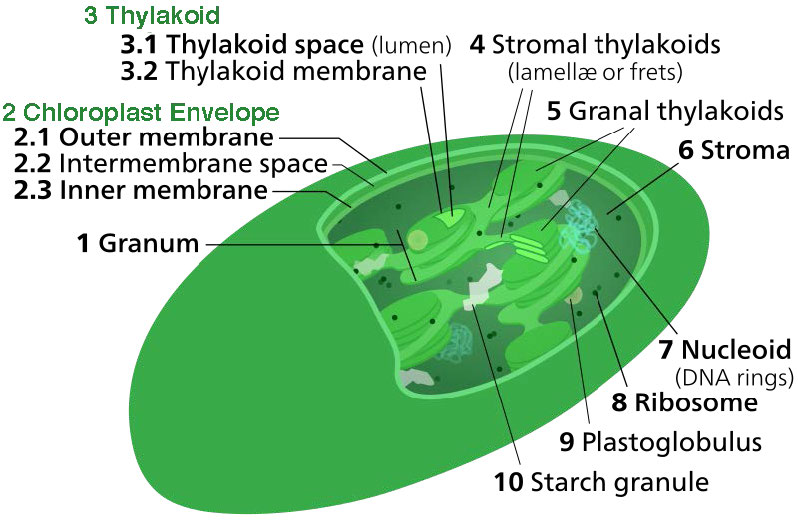
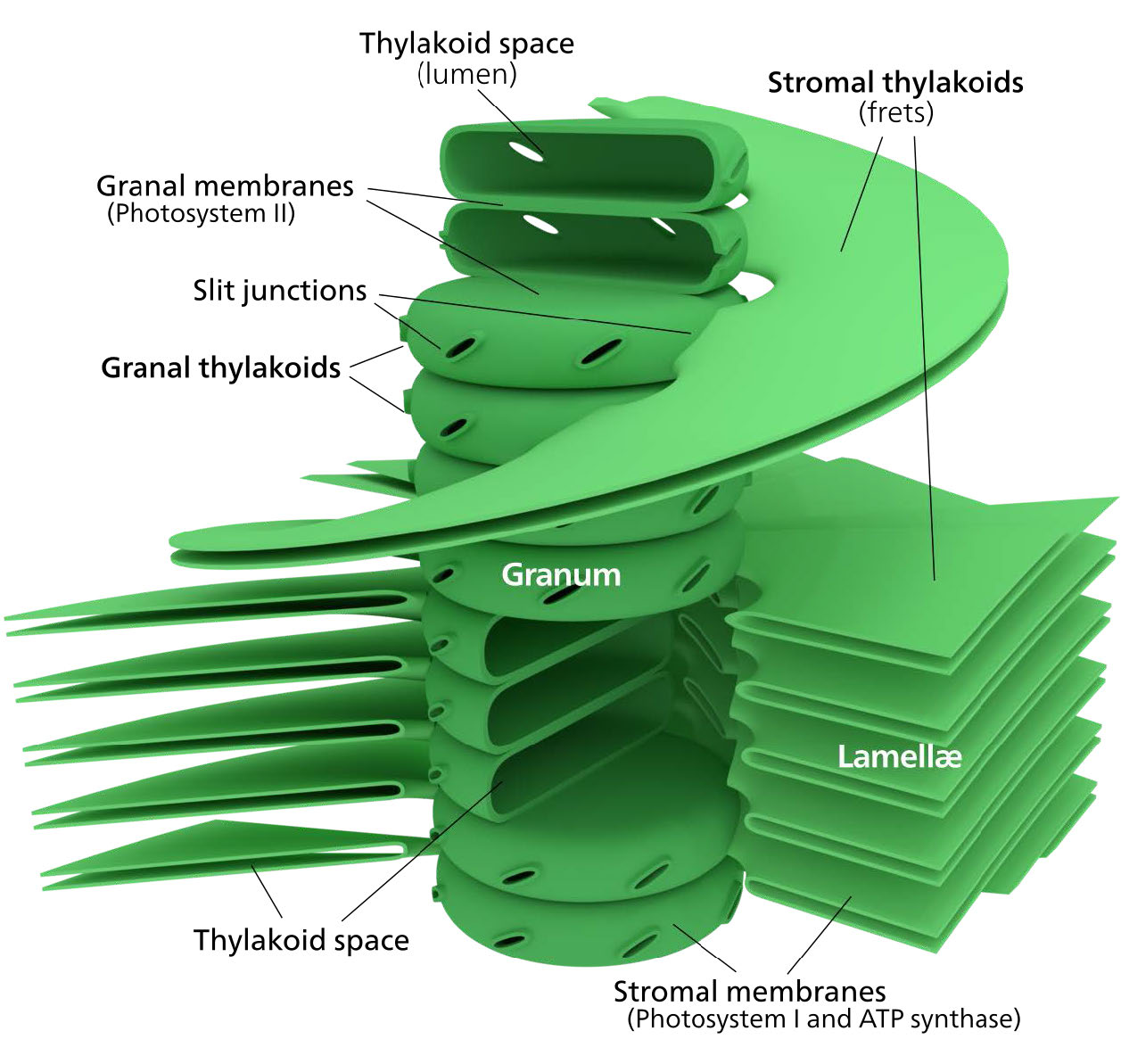
La membrana del cloroplasto tiene una membrana interna de fosfolípidos, una membrana externa de fosfolípidos y una región entre ellas llamada espacio intermembrana (Figura 5.61). Dentro de la membrana interna del cloroplasto se encuentra el estroma, en el que se ubican el ADN del cloroplasto y las enzimas del ciclo Calvino. También dentro del estroma están apilados, discos aplanados conocidos como tilacoides que se definen por sus membranas tilacoides. El espacio dentro de las membranas tilacoides se denomina espacios tilacoides o luz tilacoidea. Los complejos proteicos que contienen los pigmentos absorbentes de luz, conocidos como fotosistemas, se localizan en la membrana tilacoidea. Además de clorofilas, también están presentes carotenos y xantofilas, permitiendo la absorción de energía lumínica en un rango más amplio. Los mismos pigmentos son utilizados por las algas verdes y las plantas terrestres.
Las algas pardas y las diatomeas agregan fucoxantina (una xantofila) y las algas rojas agregan ficoeritrina a la mezcla. En plantas y algas, los pigmentos se mantienen de una manera muy organizada complejos llamados proteínas de antena que ayudan a canalizar la energía, a través de la transferencia de energía de resonancia, hacia las clorofilas del centro de reacción. Un sistema tan organizado se llama complejo de recolección de luz. Los complejos de transporte de electrones de la fotosíntesis también se localizan en las membranas tilacoides.
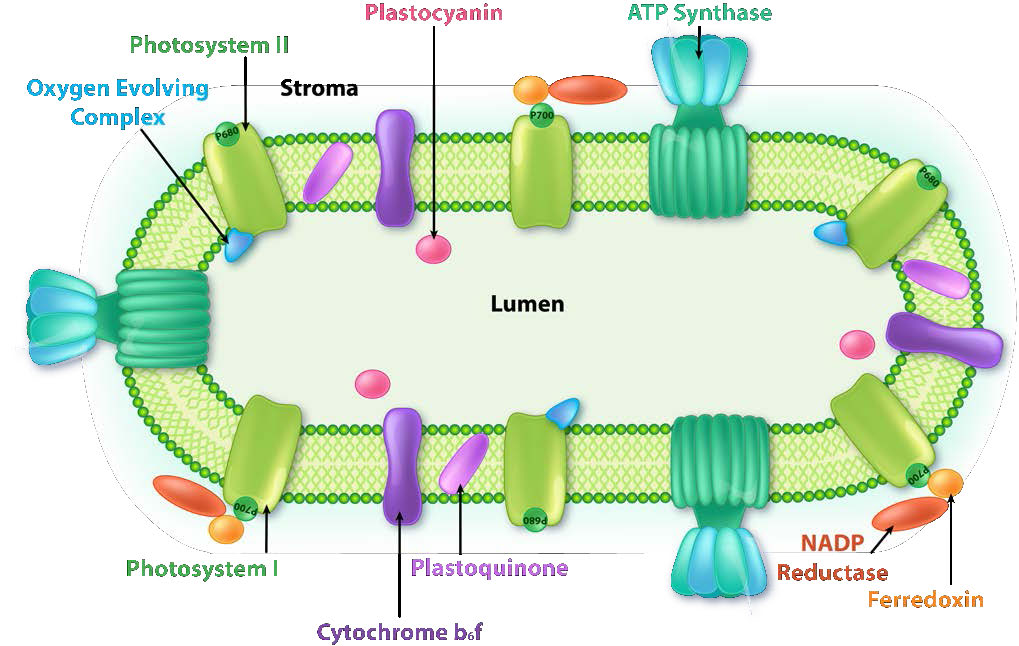
Figura\(\PageIndex{6}\): Complejos en la membrana tilacoidea. Imagen de Aleia Kim
Reacciones lumínicas de la fotosíntesis
En los cloroplastos, las reacciones lumínicas de fotosíntesis que implican transferencia de electrones ocurren en las membranas tilacoides (Figura\(\PageIndex{6}\)). Las reacciones bioquímicas separadas que implican la asimilación de dióxido de carbono para producir glucosa se conocen como el ciclo Calvino, también a veces referido como las “reacciones oscuras”. Esto se discutirá en otra parte de la sección sobre metabolismo (AQUÍ).
Los cloroplastos son donde se captura la energía de la luz, los electrones se eliminan del agua, se libera oxígeno, se produce el transporte de electrones, se forma el NADPH y se genera ATP. La membrana tilacoidea corresponde a la membrana interna de la mitocondria para el transporte de electrones y bombeo de protones (Figura\(\PageIndex{4}\)).
La membrana tilacoidea hace su magia utilizando cuatro complejos proteicos principales. Estos incluyen Fotosistema II (PS II), Complejo Citocromo b6f (CB6f), Fotosistema I (PS I) y ATP sintasa. Los papeles de estos complejos, respectivamente, son capturar energía luminosa, crear un gradiente de protones a partir del movimiento de electrones, capturar energía luminosa (nuevamente) y usar energía de gradiente de protones del proceso general para sintetizar ATP.
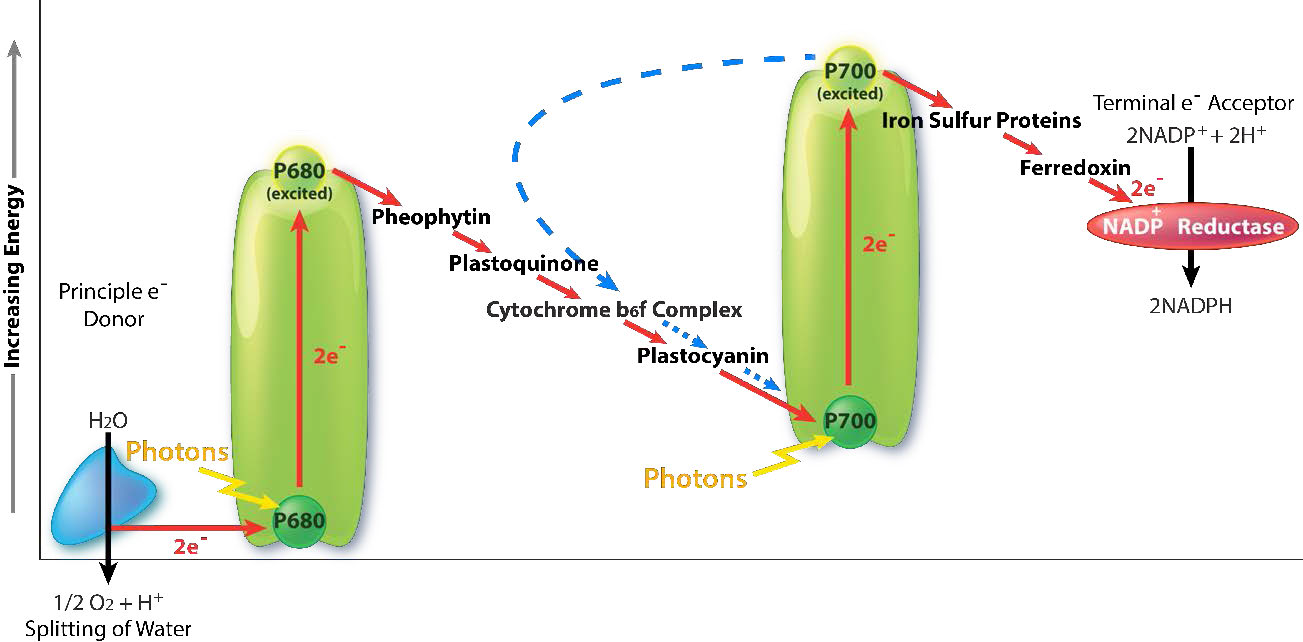
Cosecha ligera
La recolección de la energía de la luz comienza en PS II con la absorción de un fotón de luz en un centro de reacción. PS II realiza mejor este trabajo con luz a una longitud de onda de 680 nm y pierde fácilmente un electrón a la excitación cuando esto ocurre, dejando PS II con una carga positiva. Este electrón debe ser reemplazado. La última fuente de reemplazo de electrones es el agua, pero el agua debe perder cuatro electrones y PS II solo puede aceptar uno a la vez.
Centros de Manganeso
Un complejo intermedio de evolución de oxígeno (OEC) contiene cuatro centros de manganeso que proporcionan el electrón de reemplazo inmediato que el PSII requiere. Después de que cuatro electrones han sido donados por la OEC a PS II, la OEC extrae cuatro electrones de dos moléculas de agua, liberando oxígeno y arrojando cuatro protones en el espacio tilacoide, contribuyendo así al gradiente de protones. El electrón excitado de PS II debe pasarse a otro portador muy rápidamente, para que no vuelva a decairse a su estado original. Lo hace, dando su electrón en picosegundos a la feofitina (Figura\(\PageIndex{8}\)).
La feofitina pasa el electrón a plastoquinonas unidas a proteínas. El primero se conoce como PQA. PQA entrega el electrón a una segunda plastoquinona (PQB), que espera un segundo electrón y recoge dos protones para convertirse en PQH2, también conocido como plastoquinol (Figura\(\PageIndex{9}\)). PQH2 los pasa al complejo Citocromo b6f (CB6f) que utiliza el paso de electrones a través de él para bombear protones al espacio tilacoide. La ATP sintasa produce ATP a partir del gradiente de protones creado de esta manera. Cb6f deja caer el electrón en plastocianina, que lo retiene hasta que el siguiente proceso de excitación comienza con la absorción de otro fotón de luz a 700 nm por PS I.
Absorción de luz en PS I
Con la absorción de un fotón de luz por PS I, comienza un proceso, que es similar al proceso en PS II. PS I gana una carga positiva como resultado de la pérdida de un electrón excitado y aleja el electrón en plastocianina de él. En tanto, el electrón excitado de PS I pasa a través de una proteína hierro-azufre, que le da el electrón a la ferredoxina (otra proteína de azufre de hierro). La ferredoxina luego pasa el electrón a la última proteína del sistema conocido como ferredoxina:NADP+ oxidorreductasa, que le da el electrón y un protón a NADP+, creando NADPH.
Tenga en cuenta que la reducción de NADP+ a NADPH requiere dos electrones y un protón, por lo que los cuatro electrones y dos protones de la oxidación del agua darán como resultado la producción de dos moléculas de NADPH. En este punto, el ciclo de luz está completo: el agua se ha oxidado, se ha creado ATP y se ha hecho NADPH. Los electrones se han abierto camino del agua al NADPH a través de portadores en la membrana tilacoidea y su movimiento ha liberado suficiente energía para producir ATP. La energía para todo el proceso provino de cuatro fotones de luz.
Los dos fotosistemas que realizan toda esta magia son complejos proteicos que son similares en estructura y medios de operación. Absorben fotones con alta eficiencia de manera que cada vez que un pigmento en el centro de reacción fotosintético absorbe un fotón, un electrón del pigmento se excita y se transfiere a otra molécula casi instantáneamente. Esta reacción se llama separación de carga fotoinducida y es un medio único de transformar la energía lumínica en formas químicas.
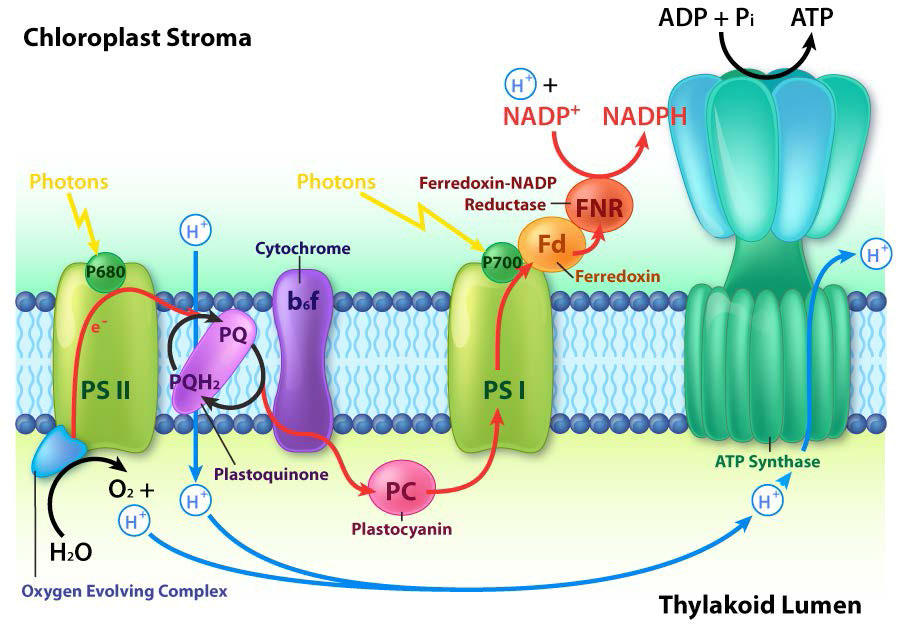
Fotofosforilación cíclica
Además del camino descrito anteriormente para el movimiento de electrones a través de PS I, las plantas tienen una ruta alternativa que los electrones pueden tomar. En lugar de que los electrones pasen a través de la ferredoxina para formar NADPH, toman un camino hacia atrás a través del complejo b6f de bombeo de protones. Este sistema, llamado fotofosforilación cíclica (Figura\(\PageIndex{8}\)) que genera más ATP y ningún NADPH, es similar a un sistema que se encuentra en bacterias verdes de azufre. La capacidad de las plantas para cambiar entre fotosistemas no cíclicos y cíclicos les permite hacer la proporción adecuada de ATP y NADPH que necesitan para la asimilación de carbono en la fase oscura de la fotosíntesis. Esta relación resulta ser de 3 ATPs a 2 NADPh.
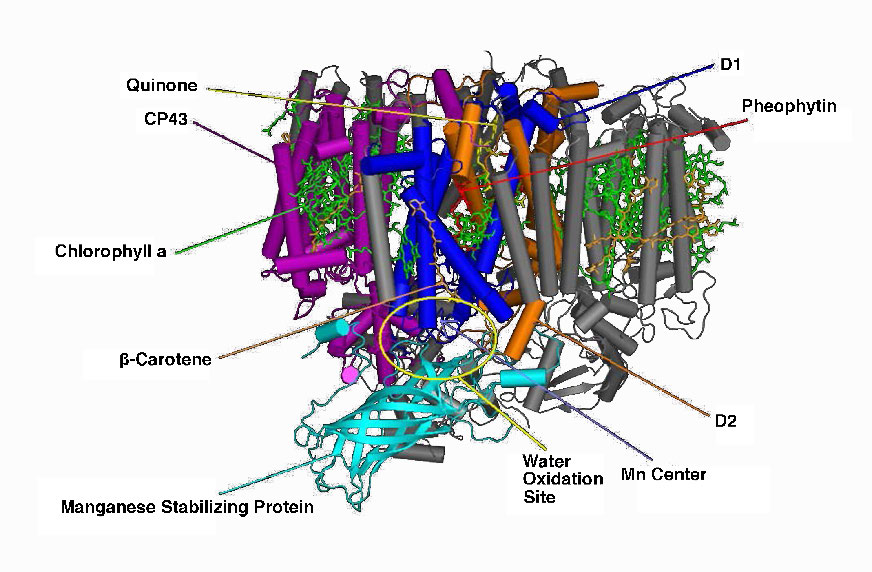
Figura\(\PageIndex{9}\) - Fotosistema II de cianobacterias. Wikipedia
Energía fotosintética
La salida de la parte de fotofosforilación de la fotosíntesis (O2, NADPH y ATP), por supuesto, no es el final del proceso de fotosíntesis. Para la planta en crecimiento, el NADPH y el ATP se utilizan para capturar el dióxido de carbono de la atmósfera y convertirlo (en última instancia) en glucosa y otros compuestos de carbono importantes. Esto, como se señaló anteriormente, ocurre en el Ciclo Calvino (ver AQUÍ) en lo que se denomina la fase oscura del proceso. El oxígeno liberado en el proceso es necesario para la respiración de todas las formas de vida aeróbicas en la Tierra. En efecto, se cree que esencialmente todo el oxígeno en la atmósfera hoy en día es el resultado de la división del agua en la fotosíntesis a lo largo de los muchos eones que ha existido el proceso.