
5.2: Transporte de electrones y fosforilación oxidativa
- Page ID
- 53047

Fuente: BiochemFFA_5_2.pdf. Todo el libro de texto está disponible de forma gratuita de los autores en http://biochem.science.oregonstate.edu/content/biochemistry-free-and-easy

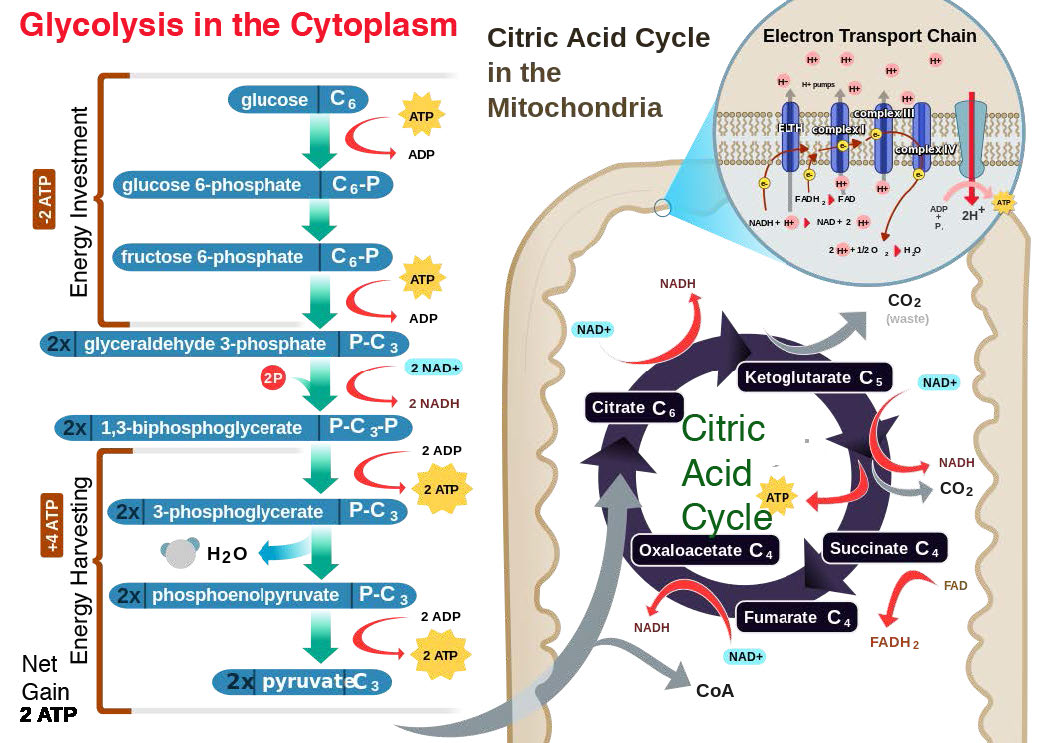
En las células eucariotas, la gran mayoría de la síntesis de ATP ocurre en las mitocondrias en un proceso llamado fosforilación oxidativa. Incluso las plantas, que generan ATP por fotofosforilación en cloroplastos, contienen mitocondrias para la síntesis de ATP a través de la fosforilación oxidativa.
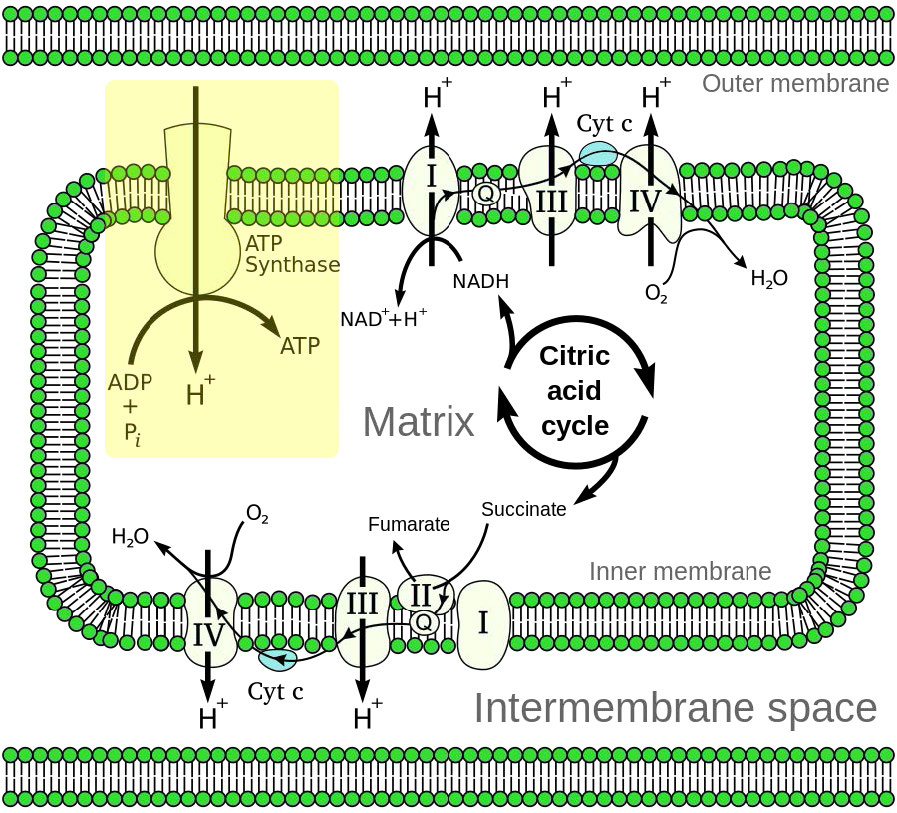
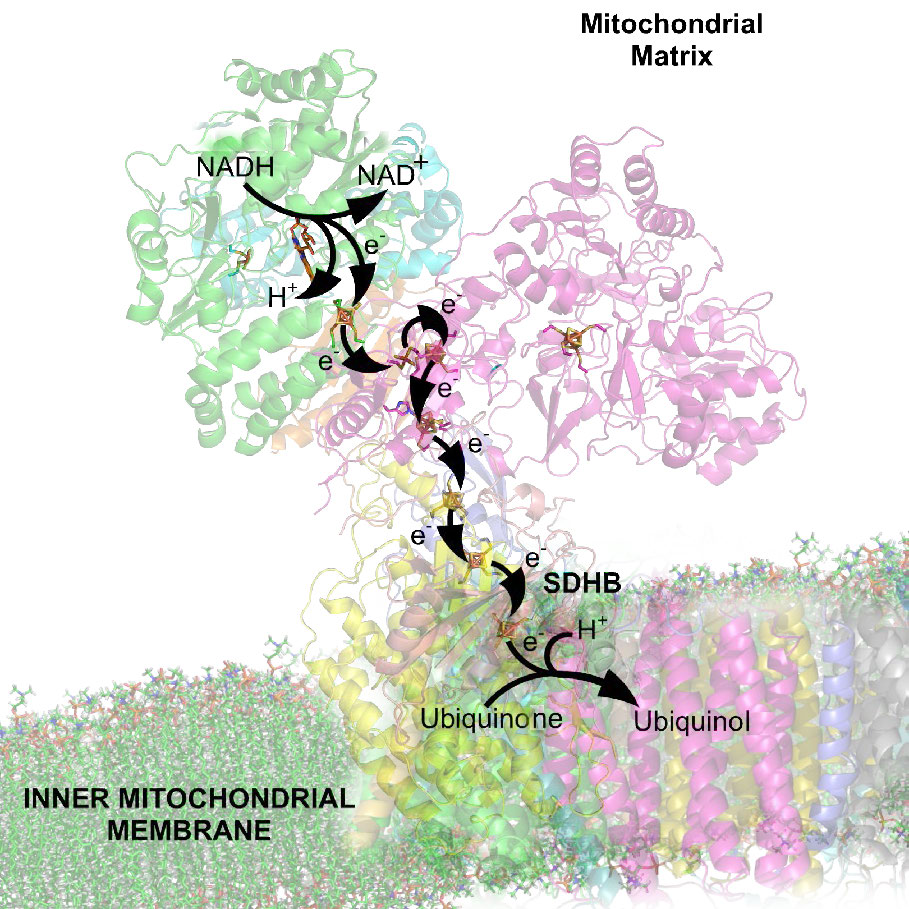
La fosforilación oxidativa está ligada a un proceso conocido como transporte de electrones (Figura 5.14). El sistema de transporte de electrones, localizado en la membrana mitocondrial interna, transfiere electrones donados por los portadores de electrones reducidos NADH y FADH2 (obtenidos de la glucólisis, el ciclo del ácido cítrico u oxidación de ácidos grasos) a través de una serie de aceptores de electrones, al oxígeno. Como veremos, el movimiento de electrones a través de complejos del sistema de transporte de electrones esencialmente “carga” una batería que se utiliza para producir ATP en la fosforilación oxidativa. De esta manera, la oxidación de azúcares y ácidos grasos se acopla a la síntesis de ATP, extrayendo efectivamente energía de los alimentos.
Modelo quimiosmótico
El Dr. Peter Mitchell introdujo una propuesta radical en 1961 para explicar el mecanismo por el cual las mitocondrias producen ATP. Se le conoce como la hipótesis quimiosmótica y se ha demostrado a lo largo de los años que es correcta. Mitchell propuso que la síntesis de ATP en las mitocondrias depende de un gradiente electroquímico, a través de la membrana interna mitocondrial, que surge en última instancia de la energía de portadores de electrones reducidos, NADH y FADH2.
Transporte de electrones
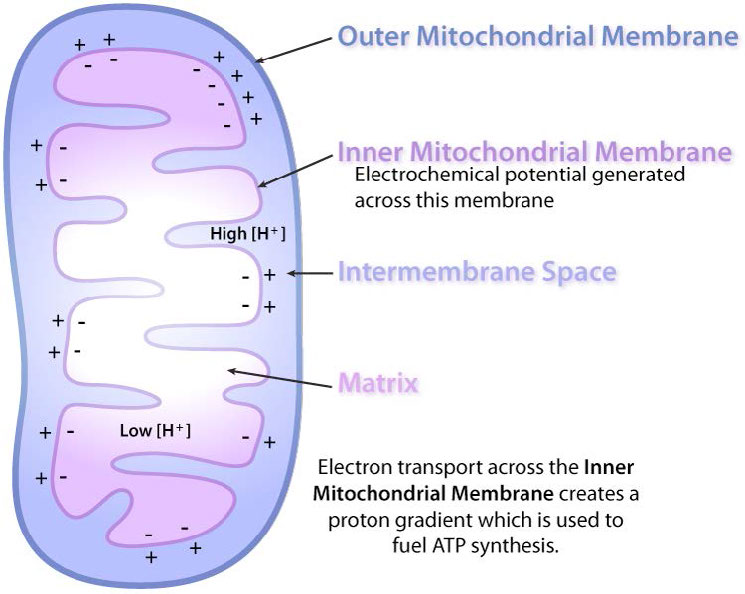
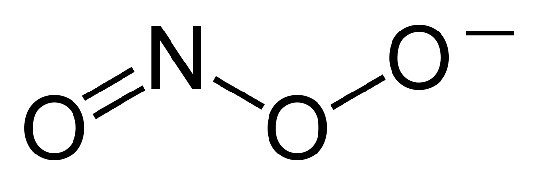
Además, la propuesta establece que el gradiente se crea cuando NADH y FADH2 transfieren sus electrones a un sistema de transporte de electrones (ETS) ubicado en la membrana mitocondrial interna. El movimiento de electrones a través de una serie de portadores de electrones se acopla al bombeo de protones fuera de la matriz mitocondrial a través de la membrana mitocondrial interna hacia el espacio entre las membranas interna y externa. El resultado es la creación de un gradiente de protones cuya energía potencial puede ser utilizada para producir ATP. Los electrones se combinan con oxígeno y protones al final del ETS para hacer agua.
ATP sintasa
En la fosforilación oxidativa, la síntesis de ATP se logra como resultado de que los protones reingresan a la matriz mitocondrial a través del complejo transmembrana ATP sintasa, que combina ADP con fosfato inorgánico para producir ATP. Central para el buen funcionamiento de las mitocondrias a través de este proceso es la presencia de una membrana interna mitocondrial intacta impermeable a los protones.
Acoplamiento apretado
Cuando este es el caso, se dice que existe un acoplamiento estrecho entre el transporte de electrones y la síntesis de ATP (llamada fosforilación oxidativa). Los químicos que permeabilizan la membrana mitocondrial interna a protones provocan el desacoplamiento, es decir, permiten que los protones vuelvan a la matriz mitocondrial, en lugar de a través de la ATP sintasa, de manera que el movimiento de electrones a través del ETS ya no está vinculado a la síntesis de ATP.
Plantas de energía
Las mitocondrias se llaman las plantas de energía de la célula porque la mayor parte del ATP de una célula se produce allí en el proceso de fosforilación oxidativa. El mecanismo por el cual se produce ATP en la fosforilación oxidativa es uno de los más interesantes en toda la biología.
Consideraciones
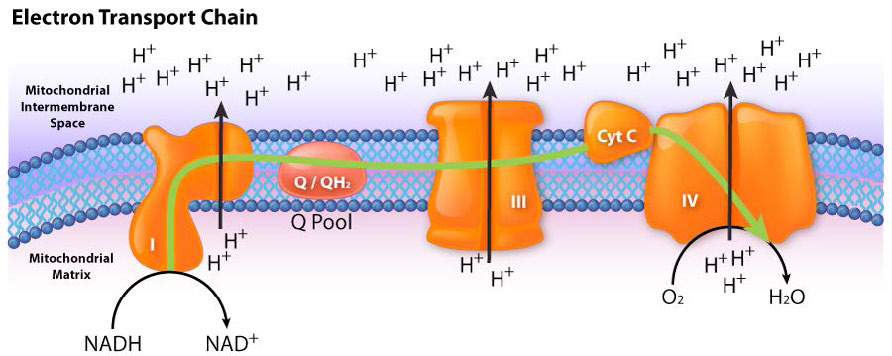
El proceso tiene tres consideraciones principales. El primero es eléctrico: los electrones de portadores de electrones reducidos, como NADH y FADH2, ingresan al sistema de transporte de electrones a través del Complejo I y II, respectivamente. Como se ve en la Figura 5.16 y la Figura 5.17, los electrones se mueven de un complejo al siguiente, no muy diferente de la forma en que se mueven a través de un circuito eléctrico. Dicho movimiento ocurre como resultado de un conjunto de reacciones de reducción-oxidación (redox) con electrones que pasan de un potencial de reducción más negativo a uno más positivo.
Se puede pensar que esto ocurre como un proceso en el que los portadores “quitan” electrones de complejos con menor potencial de reducción, de la misma manera que un matón toma el dinero del almuerzo de un niño más pequeño. En este esquema, el mayor “matón” es el oxígeno en el Complejo IV. Los electrones ganados por un portador hacen que se reduzca, mientras que el portador que cede los electrones se oxida.
Entrada de electrones al sistema
El movimiento de electrones a través de la cadena comienza ya sea por 1) transferencia del NADH al Complejo I (Figura 5.16) o 2) el movimiento de electrones a través de un FADH2 unido covalentemente (Figura 5.17) en la succinato deshidrogenasa unida a membrana (Complejo II). (Un punto de entrada alternativo para los electrones de FADH2 es la Flavoproteína de Transferencia de Electrones a través de la flavoproteína deshidrogenasa de transferencia de electrones, no mostrada).
Policía de tránsito
Tanto el Complejo I como el II pasan electrones a la coenzima Q de la membrana interna (CoQ - Figuras 5.18 y 5.19). En cada caso, la coenzima Q acepta electrones en pares y los pasa al Complejo III (CoQH2-citocromo c reductasa) individualmente. La coenzima Q actúa así como un policía de tráfico, regulando el flujo de electrones a través del ETS.
Estación de acoplamiento
El complejo III es una estación de acoplamiento o intercambio para el portador de electrones entrante (coenzima Q) y el portador saliente (citocromo c). El movimiento de electrones de la Coenzima Q al Complejo III y luego al citocromo C ocurre como resultado de lo que se conoce como el ciclo Q (ver más adelante).
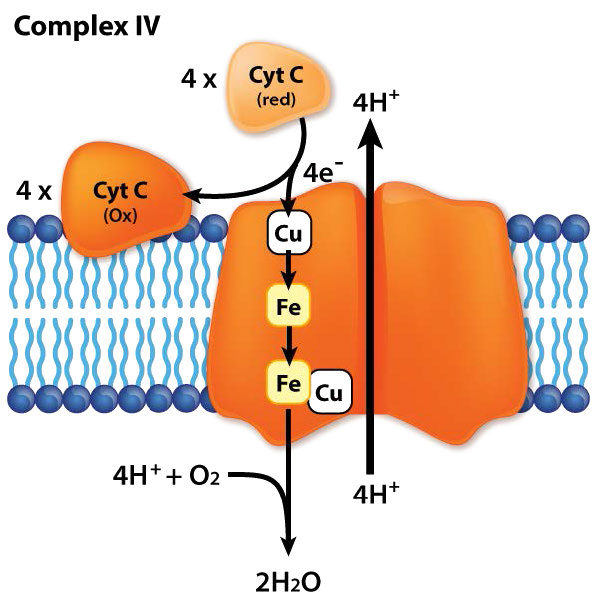
El complejo III actúa para transportar electrones de CoQ al citocromo c. El citocromo c toma un electrón del Complejo III y lo pasa al Complejo IV (citocromo oxidasa). El complejo IV es el receptor proteico final de los electrones. Los pasa a oxígeno molecular (O2) para hacer dos moléculas de agua. Hacer dos moléculas de agua requiere cuatro electrones, por lo que el Complejo IV debe aceptar, manejar y pasar al oxígeno molecular cuatro electrones separados, haciendo que el estado de oxidación del oxígeno se cambie secuencialmente con la adición de cada electrón.
Bombeo de protones
A medida que los electrones pasan a través de los complejos I, III y IV, hay una liberación de una pequeña cantidad de energía en cada paso, que se utiliza para bombear protones de la matriz mitocondrial (dentro de la mitocondria) y depositarlos en el espacio intermembrana (entre las membranas interna y externa de la mitocondria). El efecto de esta redistribución es aumentar el potencial eléctrico y químico a través de la membrana.
Energía potencial
Como se discutió anteriormente, los gradientes electroquímicos tienen energía potencial. Los estudiantes pueden pensar en el proceso como “cargar la batería”. Al igual que una batería cargada, el potencial que surge del diferencial de protones a través de la membrana se puede usar para hacer cosas. En la mitocondria, lo que hace el gradiente de protones es facilitar la producción de ATP a partir de ADP y Pi. Este proceso se conoce como fosforilación oxidativa, debido a que la fosforilación de ADP a ATP depende de las reacciones oxidativas que ocurren en las mitocondrias.
Habiendo entendido el panorama general de la síntesis de ATP ligada al movimiento de electrones a través del ETS, analizaremos más de cerca los componentes individuales del ETS.
Complejo I
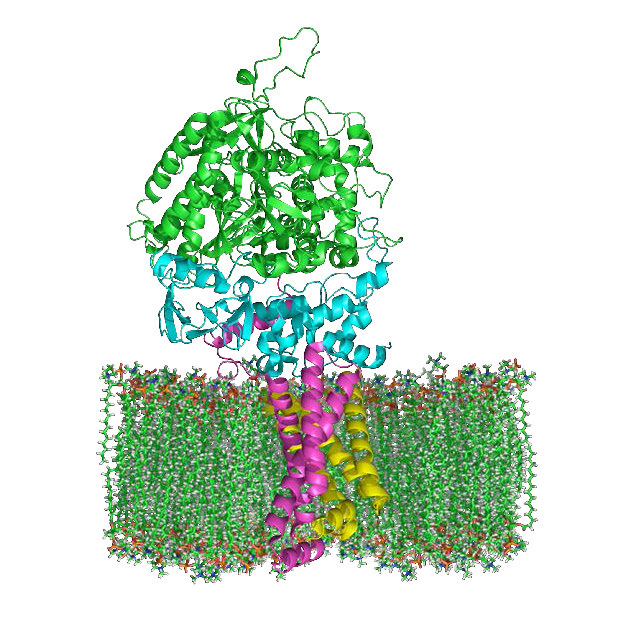
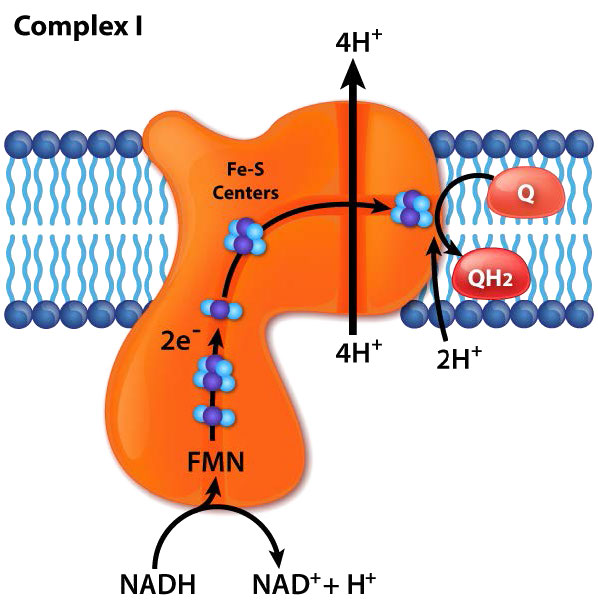
El complejo I (también llamado NADH:ubiquinona oxidorreductasa o NADH deshidrogenasa (ubiquinona)) es el aceptor de electrones de NADH en la cadena de transporte de electrones y el complejo más grande que se encuentra en ella.
El complejo I contiene 44 cadenas polipeptídicas individuales, numerosos centros de hierro-azufre, una molécula de mononucleótido de flavina (FMN) y tiene forma de L con aproximadamente 60 dominios transmembrana. En el proceso de transporte de electrones a través de ella, cuatro protones se bombean a través de la membrana interna hacia el espacio intermembrana y los electrones se mueven del NADH a la coenzima Q, convirtiéndola de ubiquinona (sin electrones) a ubiquinol (ganancia de dos electrones). Una forma intermedia, la ubisemiquinona (ganancia de un electrón), se encuentra en el ciclo Q.
Los electrones viajan a través del complejo a través de siete centros primarios de hierro y azufre. El inhibidor más conocido del complejo, la rotenona, actúa uniéndose al sitio de unión a CoQ. Otros inhibidores incluyen ADP-ribosa (se une al sitio NADH) y piericidina A (análogo de rotenona). El proceso de transferencia de electrones a través del complejo I es reversible y cuando esto ocurre, se puede generar fácilmente superóxido (una especie reactiva de oxígeno).
Complejo II
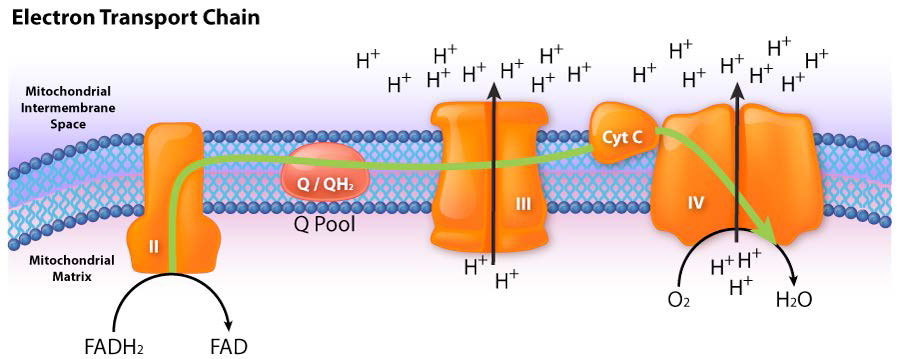
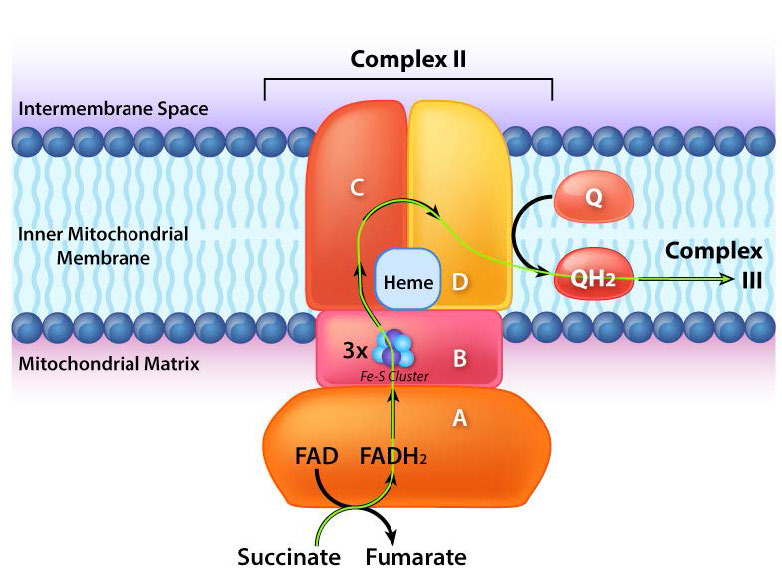
El complejo II (también llamado succinato deshidrogenasa o succinato-coenzima Q reductasa) es una enzima unida a membrana del ciclo del ácido cítrico que juega un papel en el proceso de transporte de electrones, transfiriendo electrones desde su FADH2 unido covalentemente a la coenzima Q. El proceso ocurre, como se muestra en la Figura 5.20 y Figura 5.21, con transferencia de electrones de succinato a FAD para formar FADH2 y fumarato. FADH2, a su vez, dona electrones a un sistema de retransmisión de grupos hierro-azufre y finalmente reducen la ubiquinona (CoQ) junto con dos protones de la matriz a ubiquinol. El papel del grupo hemo en el proceso no está claro. Los inhibidores del proceso incluyen carboxina, malonato, malato y oxaloacetato. Se cree que el papel de los intermedios del ciclo del ácido cítrico como inhibidores se debe a la inhibición de la inversión del proceso de transferencia que puede producir superóxido.
Coenzima Q
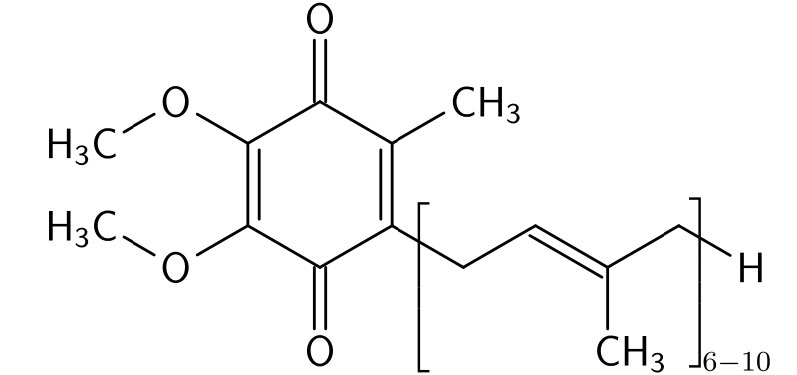
La coenzima Q (Figura 5.23) es una 1,4 benzoquinona cuyo nombre suele darse como Coenzima Q10, CoQ o Q10. El 10 en el nombre se refiere al número de unidades de isoprenilo que contiene que lo anclan a la membrana interna mitocondrial. CoQ es una sustancia lipídica similar a las vitaminas que se encuentra en la mayoría de las células eucariotas como componente del sistema de transporte de electrones. El requerimiento de CoQ aumenta con el aumento de las necesidades energéticas de las células, por lo que las concentraciones más altas de CoQ en el cuerpo se encuentran en los tejidos que son los más activos metabólicamente: corazón, hígado y riñón.
Tres formas
CoQ es útil por su capacidad para transportar y donar electrones y particularmente porque puede existir en formas con dos electrones extra (completamente reducidos - ubiquinol), un electrón extra (semi-reducido - ubisemiquinona), o sin electrones adicionales (completamente oxidado - ubiquinona). Esta capacidad permite a CoQ proporcionar transición entre la primera parte del sistema de transporte de electrones que mueve electrones en pares y la última parte del sistema que mueve electrones uno a la vez.
Complejo III
El complejo III (también conocido como coenzima Q: citocromo c — oxidorreductasa o el complejo citocromo bc1 - Figura 5.24) es el tercer complejo aceptor de electrones del sistema de transporte de electrones. Es una proteína transmembrana con múltiples subunidades presentes en las mitocondrias de todos los organismos eucariotas aeróbicos y en la membrana celular de casi todas las bacterias. El complejo contiene 11 subunidades, una ferredoxina 2-hierro, citocromos b y c1 y pertenece a la familia de enzimas oxidorreductasa.
Acepta electrones de la coenzima Q en el transporte de electrones y los pasa al citocromo c. En este ciclo, conocido como el ciclo Q, los electrones llegan de CoQ en pares, pero pasan al citocromo c individualmente. En el proceso general, se consumen dos protones de la matriz y cuatro protones se bombean al espacio intermembrana. El movimiento de electrones a través del complejo puede ser inhibido por la antimicina A, mixotiazol y estigmatelina. El complejo III también está implicado en la creación de superóxido (una especie reactiva de oxígeno) cuando los electrones del mismo se escapan de la cadena de transferencia. El fenómeno es más pronunciado cuando la antimicina A está presente.
Ciclo Q
En el ciclo Q, los electrones se pasan del ubiquinol (QH2) al citocromo c usando el Complejo III como estación de acoplamiento intermedia para la transferencia. Dos pares de electrones entran desde QH2 y un par es devuelto a otro CoQ para volver a hacer QH2. El otro par se dona por separado a dos moléculas diferentes de citocromo c.
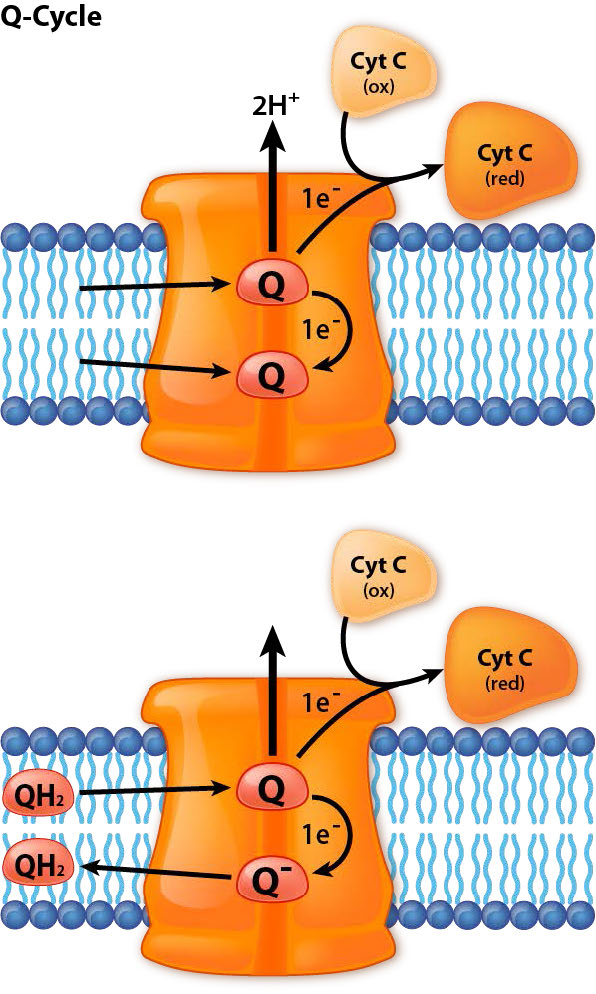
Paso uno
El ciclo Q ocurre en un proceso de dos pasos. Primero, un muelle de ubiquinol (CoQH2) y una ubiquinona (CoQ) en el Complejo III. El ubiquinol transfiere dos electrones al Complejo III. Un electrón va a un citocromo c acoplado, reduciéndolo y sale (reemplazado por un citocromo c oxidado). El otro va a la uniquinona acoplada para crear la semiubiquinona semirreducida (CoQ.-) y dejando atrás una ubiquinona, que sale. Este es el final del paso 1.
Paso dos
El hueco dejado por la ubiquinona (Q) que partió es reemplazado por otro ubiquinol (QH2). También dona dos electrones al Complejo III, que los divide. Uno va al citocromo c oxidado recién acoplado, que se reduce y sale. El otro va a la ubisemiquinona. Dos protones de la matriz se combinan con ella para hacer otro ubiquinol. Ella y la ubiquinona creada por la donación de electrones salen del Complejo III y el proceso comienza de nuevo. En el proceso general, se consumen dos protones de la matriz y cuatro protones se bombean al espacio intermembrana.
Citocromo c
El citocromo c (Figura 5.26) es una proteína pequeña (12,000 Daltons), altamente conservada, desde especies unicelulares hasta animales, que se asocia libremente con la membrana mitocondrial interna donde funciona en el transporte de electrones. Contiene un grupo hemo que se utiliza para transportar un solo electrón del Complejo III al Complejo IV. El citocromo c también juega un papel importante en la apoptosis en organismos superiores. El daño a la mitocondria que resulta en la liberación del citocromo c puede estimular el ensamblaje del apoptosoma y la activación de la cascada de caspasa que conduce a la muerte celular programada.
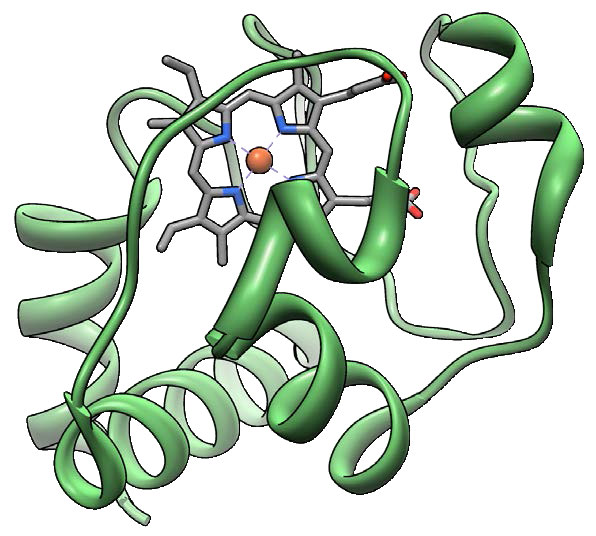
Complejo IV
El complejo IV, también conocido como citocromo c oxidasa, es una proteína de membrana integral de 14 subunidades al final de la cadena de transporte de electrones (Figura 5.27). Se encarga de aceptar un electrón cada una de cuatro proteínas del citocromo c y agregarlas al oxígeno molecular (O2) junto con cuatro protones de la matriz mitocondrial para formar dos moléculas de agua. Cuatro protones de la matriz también se bombean en el espacio intermembrana en el proceso. El complejo tiene dos moléculas de hemo, dos citocromos (a y a3) y dos centros de cobre (llamados CuA ad CuB). El citocromo c atraca cerca del CuA y le dona un electrón. El CuA reducido pasa el electrón al citocromo a, que lo vuelve al centro A3-CuB donde se reduce el oxígeno. Se cree que los cuatro electrones pasan a través del complejo rápidamente dando como resultado la reducción completa de la molécula de oxígeno-oxígeno sin la formación de un peróxido intermedio o superóxido, en contraste con las predicciones anteriores.
Respirasoma
Se ha especulado desde hace muchos años que un supercomplejo de portadores de electrones en la membrana interna de la mitocondria puede existir en células con portadores individuales haciendo contacto físico entre sí. Esto haría que las reacciones de transferencia fueran más eficientes, minimizarían la producción de especies reactivas de oxígeno y serían similares a los metabolones de las enzimas de la vía metabólica, para lo cual hay alguna evidencia. Ahora, parece estar acumulando evidencia de que los complejos I, III y IV forman un supercomplejo, al que se le ha denominado respirasome1.
Fosforilación oxidativa
El proceso de fosforilación oxidativa utiliza la energía del gradiente de protones establecido por el sistema de transporte de electrones como medio de fosforilación de ADP para producir ATP. El establecimiento del gradiente de protones depende del transporte de electrones. Si el transporte de electrones se detiene o si se compromete la impermeabilidad de la membrana mitocondrial interna a los protones, la fosforilación oxidativa no ocurrirá porque sin el gradiente de protones para impulsar la ATP sintasa, no habrá síntesis de ATP.
ATP sintasa
El complejo proteico que recoge energía del gradiente de protones y lo utiliza para producir ATP a partir de ADP es una enzima que tiene varios nombres: Complejo V, PTAS (Proton Translocating ATP Synthase) y ATP sintasa (Figura 5.29). Central para su función es el movimiento de protones a través de él (desde el espacio intermembrana de vuelta a la matriz). Los protones solo proporcionarán energía para producir ATP si su concentración es mayor en el espacio intermembrana que en la matriz y si hay ADP disponible.
Es posible, en algunos casos, que la concentración de protones sea mayor dentro de la matriz que fuera de ella. Cuando esto sucede, la ATP sintasa puede correr hacia atrás, con protones moviéndose de adentro hacia afuera, acompañados de la conversión de ATP a ADP + Pi. Esta no suele ser una circunstancia deseable y existen algunos controles para reducir su ocurrencia.
Normalmente, la concentración de ATP será mayor dentro de la mitocondria y la concentración de ADP será mayor fuera de la mitocondria. Sin embargo, cuando la tasa de síntesis de ATP excede la tasa de uso de ATP, entonces las concentraciones de ATP aumentan fuera de la mitocondria y las concentraciones de ADP caen en todas partes.
Esto puede suceder, por ejemplo, durante periodos de descanso. Tiene el efecto general de reducir el transporte y así disminuir la concentración de ADP dentro de la matriz. La reducción de la concentración de ADP en la matriz reduce la fosforilación oxidativa y tiene efectos sobre el control respiratorio (ver AQUÍ).
Otra consideración importante es que cuando el ATP se hace en la fosforilación oxidativa, se libera en la matriz mitocondrial, pero debe transportarse al citosol para satisfacer las necesidades energéticas del resto de la célula. Esto se logra mediante la acción de la adenina nucleótido translocasa, un antipuerto que mueve ATP fuera de la matriz a cambio de ADP entrando en la matriz. Este sistema de transporte es impulsado por las concentraciones de ADP y ATP y asegura que los niveles de ADP se mantengan dentro de la mitocondria, permitiendo una síntesis continua de ATP.
Un último requisito para la síntesis de ATP a partir de ADP es que el fosfato también debe importarse a la matriz. Esto se logra mediante la acción de la translocasa de fosfato, que es un simporte que mueve el fosfato a la matriz mitocondrial junto con un protón.
Existe evidencia de que las dos translocasas y ATP sintasa pueden existir en un complejo, el cual ha sido denominado ATP sintasoma.
En resumen, el sistema de transporte de electrones carga la batería para la fosforilación oxidativa bombeando protones fuera de la mitocondria. La membrana interna intacta de la mitocondria mantiene los protones fuera, excepto aquellos que vuelven a entrar a través de ATP Sintasa. La ATP Sintasa permite que los protones vuelvan a entrar en la matriz mitocondrial y cosechar su energía para producir ATP.
Mecanismo de ATP sintasa
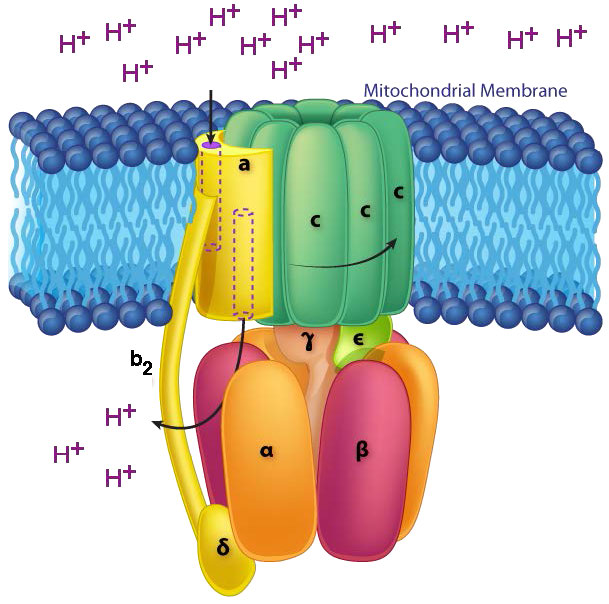
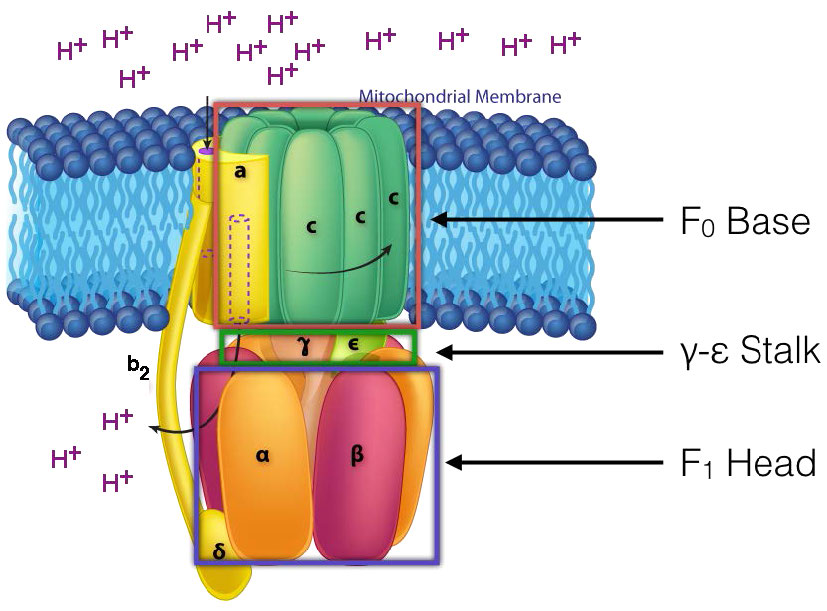
En la ATP Sintasa, los componentes giratorios, o rotor, son la porción de membrana (anillo c) de la base F0 y el tallo γ-ε, que está conectado a ella. El tallo γ-ε se proyecta en la cabeza F1 de la estructura del hongo. La cabeza F1 contiene la capacidad catalítica para producir ATP. La cabeza F1 es de estructura hexamérica con proteínas α y β emparejadas dispuestas en un trímero de dímeros. La síntesis de ATP ocurre dentro de las subunidades β.
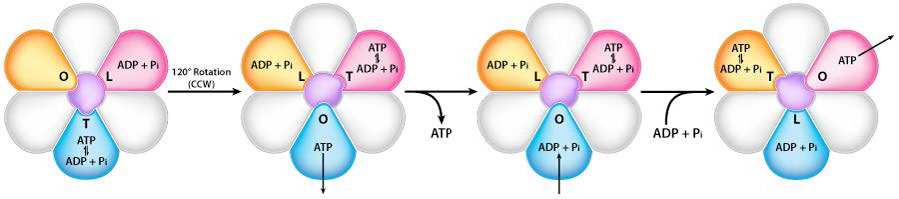
Rotación de la unidad γ
El giro del eje γ (causado por el flujo de protones) dentro del trímero α-β de la cabeza F1 hace que cada conjunto de proteínas β cambie la estructura ligeramente en tres formas diferentes llamadas Suelta, Estrecha y Abierta (L, T, O - Figura 5.31). Cada una de estas formas tiene una función.
La forma suelta se une a ADP + Pi. La forma apretada las “aprieta” juntas para formar la ATP. La forma abierta libera el ATP en la matriz mitocondrial. Así, como resultado del flujo protónico a través de la ATP sintasa, desde el espacio intermembrana hacia la matriz, el ATP se elabora a partir de ADP y Pi.
Control respiratorio
Cuando una mitocondria tiene una membrana interna intacta y los protones solo pueden regresar a la matriz al pasar a través de la ATP sintasa, se dice que los procesos de transporte de electrones y fosforilación oxidativa están estrechamente acoplados.
Interdependencia
En términos simples, acoplamiento estrecho significa que los procesos de transporte de electrones y fosforilación oxidativa son interdependientes. Sin el transporte de electrones en la célula, la fosforilación oxidativa pronto se detendrá.
Lo contrario también es cierto, porque si la fosforilación oxidativa se detiene, el gradiente de protones no se disipará ya que está siendo construido por el sistema de transporte de electrones y crecerá cada vez más. Cuanto mayor sea el gradiente, mayor será la energía necesaria para bombear protones fuera de la mitocondria. Eventualmente, si nada alivia el gradiente, éste se vuelve demasiado grande y la energía del transporte de electrones es insuficiente para realizar el bombeo. Cuando el bombeo se detiene, también lo hace el transporte de electrones.
Dependencia ADP
Otro punto relevante es que la ATP sintasa depende totalmente de un suministro de ADP. En ausencia de ADP, la ATP sintasa deja de funcionar y cuando se detiene, también lo hace el movimiento de protones de regreso a la mitocondria. Con esta información, es posible comprender el vínculo entre el uso de energía y el metabolismo. La raíz de esto, como se señaló, es el control respiratorio.
En reposo
Para ilustrar estos vínculos, consideremos primero a una persona, inicialmente en reposo, que luego de repente salta y huye. Al principio, los niveles de ATP de la persona son altos y los niveles de ADP son bajos (sin ejercicio para quemar ATP), por lo que se está produciendo poca fosforilación oxidativa y por lo tanto el gradiente de protones es alto. El transporte de electrones se mueve lentamente, en todo caso, por lo que no está usando oxígeno y la respiración de la persona es lenta, como resultado.
Ejercicio
Cuando se inicia la carrera, la contracción muscular, que utiliza energía, hace que el ATP se convierta en ADP. El aumento del ADP en las células musculares favorece la fosforilación oxidativa para intentar compensar el ATP que se quema. La ATP sintasa comienza a funcionar y los protones comienzan a regresar a la matriz mitocondrial. El gradiente de protones disminuye, por lo que se reinicia el transporte de electrones.
El transporte de electrones necesita un aceptor de electrones, por lo que el uso de oxígeno aumenta y cuando aumenta el uso de oxígeno, la persona comienza a respirar más intensamente para abastecerlo. Cuando la persona deja de correr, las concentraciones de ATP son reconstruidas por ATP sintasa. Eventualmente, cuando los niveles de ATP están completamente restaurados, los niveles de ADP disminuyen y la ATP sintasa se detiene o ralentiza considerablemente. Con poco o ningún movimiento de protones, el transporte de electrones se detiene porque el gradiente de protones es demasiado grande. Cuando el transporte de electrones se detiene, el uso de oxígeno disminuye y la velocidad de respiración se ralentiza.
El transporte de electrones es crítico
Los vínculos realmente interesantes con el metabolismo ocurren en relación con si se está produciendo o no el transporte de electrones. A partir de los ejemplos, podemos ver que el transporte de electrones se ralentizará relativamente cuando no se haga ejercicio y más rápido cuando se produzca el ejercicio (u otro uso de ATP). Recuerde que el transporte de electrones es la forma en que los portadores de electrones reducidos, NADH y FADH2, donan sus electrones al ETS, quedando oxidados a NAD+ y FAD, respectivamente.
Los portadores oxidados, como el NAD+ y el FAD, son necesarios por vías catabólicas, como la glucólisis, el ciclo del ácido cítrico y la oxidación de ácidos grasos. Las vías anabólicas, como la síntesis de ácidos grasos/grasas y la gluconeogénesis, se basan en portadores de electrones reducidos, como FADH2, NADH y el portador relacionado, NADPH.
Enlaces al ejercicio
Los altos niveles de NADH y FADH2 impiden que las vías catabólicas operen, ya que los niveles de NAD+ y FAD serán bajos y estos son necesarios para aceptar los electrones liberados durante el catabolismo por los procesos oxidativos.
Gracias al control respiratorio, cuando uno está haciendo ejercicio, los niveles de NAD+ y FAD aumentan (porque el transporte de electrones está funcionando), por lo que las vías catabólicas que necesitan NAD+ y FAD pueden funcionar. Los electrones perdidos en las reacciones de oxidación del catabolismo son capturados por NAD+ y FAD para producir NADH y FADH2, que luego suministran electrones al sistema de transporte de electrones y fosforilación oxidativa para hacer ATP más necesario.
Así, durante el ejercicio, las células se mueven a un modo de ciclado rápido entre portadores de electrones reducidos (NADH/FADH2) y portadores de electrones oxidados (NAD+/FAD). Esto permite metabolizar rápidamente los tejidos para transferir electrones a NAD+/FAD y permite que los portadores de electrones reducidos se oxiden rápidamente, permitiendo que la célula produzca ATP.
Resto
Cuando el ejercicio se detiene, los niveles de NADH y FADH2 suben (porque el transporte de electrones se está ralentizando) provocando que las vías catabólicas se ralenticen o Si uno no tiene la cantidad adecuada de ejercicio, los portadores reducidos permanecen altos en concentración durante largos períodos de tiempo. Esto significa que tenemos un exceso de energía y luego se favorecen las vías anabólicas, particularmente la síntesis de ácidos grasos, por lo que engordamos más.
Alterar el control respiratorio
Uno podría sospechar que alterar el control respiratorio podría tener algunas consecuencias muy graves y eso sería correcto. Las alteraciones pueden tomar la forma de inhibir el transporte de electrones/fosforilación oxidativa o desacoplar los dos. Estas alteraciones se pueden lograr usando compuestos con efectos específicos sobre componentes particulares del sistema.
Todos los productos químicos descritos aquí son herramientas de laboratorio y nunca deben ser utilizados por personas. El primer grupo de discusión son los inhibidores. En las mitocondrias estrechamente acopladas, inhibir el transporte de electrones o la fosforilación oxidativa también tiene el efecto de inhibir a la otra.
Inhibidores del transporte electrónico
Los inhibidores comunes del transporte de electrones incluyen rotenona y amital, que detienen el movimiento de electrones más allá del Complejo I, malonato, malato y oxaloacetato, que inhiben el movimiento de electrones a través del Complejo II, antimicina A que detiene el movimiento de electrones más allá del Complejo III, y cianuro, monóxido de carbono, azida y sulfuro de hidrógeno, que inhiben el movimiento de electrones a través del Complejo IV (Figura 5.33). Todos estos compuestos pueden detener el transporte de electrones directamente (sin movimiento de electrones) y la fosforilación oxidativa indirectamente (el gradiente de protones se disipará). Si bien algunos de estos compuestos no son comúnmente conocidos, casi todos son conscientes de los peligros del monóxido de carbono y el cianuro, los cuales pueden ser letales.
Inhibidor de ATP sintasa
También es posible utilizar un inhibidor de ATP sintasa para detener la fosforilación oxidativa directamente (sin producción de ATP) y el transporte de electrones indirectamente (gradiente de protones no aliviado por lo que se vuelve cada vez más difícil bombear protones fuera de la matriz). La oligomicina A (Figura 5.34) es un inhibidor de la ATP sintasa.
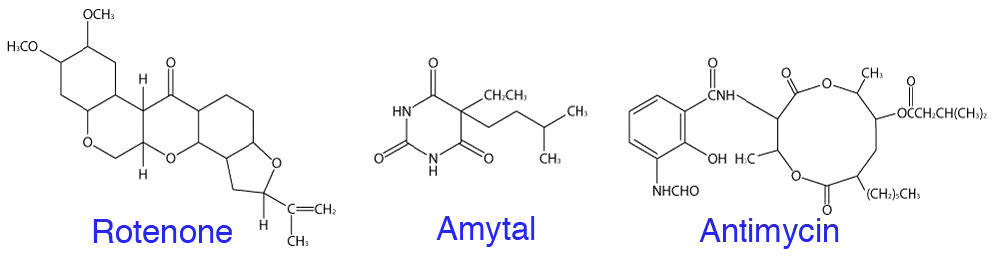
Rotenona
La rotenona, que es un producto vegetal, se utiliza como insecticida natural que está permitido para la agricultura ecológica. Cuando las mitocondrias sean tratadas con esto, el transporte de electrones se detendrá en el Complejo I y también lo hará el bombeo de protones fuera de la matriz. Cuando esto ocurre, el gradiente de protones se disipa rápidamente, deteniendo como consecuencia la fosforilación oxidativa. Existen otros puntos de entrada para los electrones además del Complejo I, por lo que este tipo de inhibición no es tan grave como el uso de inhibidores del Complejo IV, ya que no se dispone de una ruta alternativa para los electrones. Es por esta razón que el cianuro, por ejemplo, es tan venenoso.
2,4-DNP
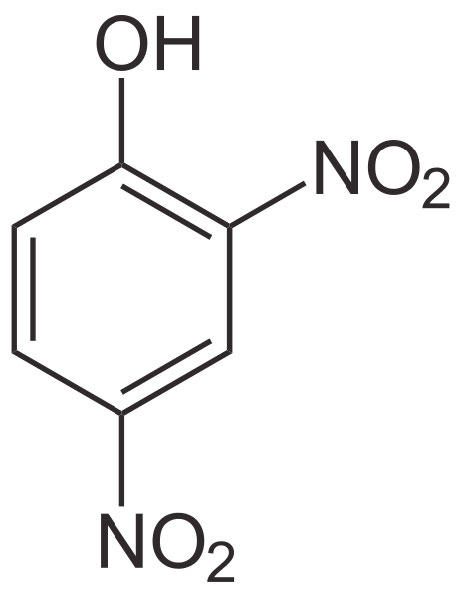
Imagina una presa que retiene el agua con una turbina que genera electricidad a través de la cual debe fluir el agua. Cuando toda el agua fluye a través de la turbina, se puede generar la cantidad máxima de electricidad. Sin embargo, si uno hace un agujero en la presa, el agua fluirá a través del agujero y se creará menos electricidad. La generación de electricidad quedará así desacoplada del flujo de agua. Si el agujero es lo suficientemente grande, el agua se drenará a través del agujero y no se hará electricidad.
Sin pasar por ATP sintasa
Imagínese, ahora, que el gradiente de protones es el equivalente del agua, la membrana interna es el equivalente de la presa y la ATP sintasa es la turbina. Cuando los protones tienen una ruta alternativa, poco o ningún ATP se hará porque los protones pasarán a través de los agujeros de la membrana en lugar de hacer girar la turbina de ATP sintasa.
Sin embargo, es importante reconocer que el desacoplamiento por 2,4 DNP funciona de manera diferente a los inhibidores del transporte de electrones o al inhibidor de la ATP sintasa. En esas situaciones, detener la fosforilación oxidativa resultó en detener indirectamente el transporte de electrones, ya que los dos procesos se acoplaron y los inhibidores no los desacoplaron. De manera similar, detener el transporte de electrones detuvo indirectamente la fosforilación oxidativa por la misma razón.
Tal no es el caso con 2,4 DNP. Detener la fosforilación oxidativa destruyendo el gradiente de protones permite que el transporte de electrones continúe sin disminuir (en realidad lo estimula), ya que el gradiente de protones no puede construirse por mucho transporte de electrones que se ejecute. En consecuencia, el transporte de electrones corre como loco pero la fosforilación oxidativa se detiene. Cuando eso sucede, los niveles de NAD+ y FAD aumentan, y las vías catabólicas funcionan sin disminuir con abundantes suministros de estos aceptores de electrones. La razón por la que tal escenario es peligroso es porque el cuerpo está utilizando todos sus recursos nutritivos, pero no se está fabricando ATP. La falta de ATP conduce a la muerte celular (y orgánica). Además, las grandes cantidades de calor generado pueden elevar la temperatura del cuerpo a niveles inseguros.

Termogenina
Uno de los subproductos del desacoplamiento del transporte de electrones es la producción de calor. Cuanto más rápido se ejecutan las vías metabólicas, más calor se genera como subproducto. Dado que 2,4 DNP hace que el metabolismo se acelere, se puede producir una cantidad considerable de calor. El desacoplamiento controlado es realmente utilizado por el cuerpo en tejidos especiales llamados grasa marrón. En este caso, las células grasas pardas utilizan el calor creado para ayudar a termorregular la temperatura de los recién nacidos.
La permeabilización de la membrana interna se logra en la grasa marrón mediante la síntesis de una proteína llamada termogenina (también conocida como proteína de desacoplamiento). La termogenina se une a la membrana interna y permite que los protones pasen a través de ella, evitando así la ATP sintasa. Como se señaló para 2,4 DNP, esto da como resultado la activación de las vías catabólicas y cuanto más catabolismo ocurre, más calor se genera.
Droga peligrosa
En el desacoplamiento, ya sea a través de la acción de una proteína desacoplante endógena o DNP, la energía que normalmente habría sido capturada en ATP se pierde como calor. En el caso del desacoplamiento por termogenina, esto sirve para el importante propósito de mantener calientes a los recién nacidos. Pero en los adultos, el desacoplamiento simplemente desperdicia la energía que se habría cosechado como ATP. Es decir, imita el hambre, a pesar de que hay mucha comida, porque la energía se disipa como calor.
Este hecho, y el aumento asociado en la tasa metabólica, llevaron a que el DNP se usara como medicamento para bajar de peso en la década de 1930. Se promocionó como una forma sin esfuerzo de perder peso sin tener que comer menos o hacer más ejercicio, fue aclamada como una píldora mágica para perder peso. Rápidamente se hizo evidente, sin embargo, que esto era muy peligroso. Muchas personas murieron por usar este medicamento antes de que se aprobaron leyes para prohibir el uso de DNP como ayuda para perder peso.
Oxidasa alternativa
Otro enfoque para generar calor que no implica romper el control respiratorio es tomado por algunos hongos, plantas y protozoos. Utilizan un transporte de electrones alternativo. En estos organismos existe una enzima llamada oxidasa alternativa (Figura 5.36). La oxidasa alternativa es capaz de aceptar electrones de CoQ y pasarlos directamente al oxígeno.
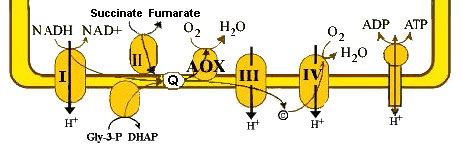
El proceso ocurre en las mitocondrias acopladas. Su mecanismo de acción es reducir el rendimiento de ATP, ya que se están bombeando menos protones por cada portador de electrones reducido. Así, las concentraciones de NAD+ aumentan, el consumo de oxígeno aumenta y la eficiencia de la producción de ATP disminuye.
Los organismos que utilizan este método deben activar las vías catabólicas por el aumento de la concentración de NAD+. Esto, a su vez produce cantidades de NADH y FADH2 necesarias para hacer cantidades suficientes de ATP. El subproducto de este aumento del catabolismo es más calor. No es sorprendente que la ruta alternativa de oxidasa pueda activarse por temperaturas frías.
Eficiencia Energética
Las células no son 100% eficientes en el uso de energía. Nada de lo que sabemos lo es. En consecuencia, las células no obtienen tanta energía de los procesos catabólicos como la ponen en los procesos anabólicos. Un buen ejemplo es la síntesis y descomposición de la glucosa, algo que suelen hacer las células hepáticas. La conversión completa de glucosa a piruvato en glucólisis (catabolismo) produce dos piruvatos más 2 NADH más 2 ATPs. Por el contrario, la conversión completa de dos piruvatos en glucosa por gluconeogénesis (anabolismo) requiere 4 ATP, 2 NADH y 2 GTP. Dado que la energía del GTP es esencialmente igual a la del ATP, la gluconeogénesis requiere una red de 4 ATP más que los rendimientos de glucólisis. Esta diferencia debe ser maquillada para que el organismo pueda satisfacer sus necesidades energéticas. Es por esta razón que comemos. Además, la ineficiencia de nuestra captura de energía en las reacciones da como resultado la producción de calor y ayuda a mantenernos calientes, como se señaló. Puedes leer más sobre la glucólisis (AQUÍ) y la gluconeogénesis (AQUÍ).
Controles metabólicos de la energía
También es de destacar que las células no suelen tener procesos tanto catabólicos como anabólicos para las mismas moléculas que ocurren simultáneamente dentro de ellas (por ejemplo, descomposición de glucosa y síntesis de glucosa) porque la célula no vería ninguna producción neta de nada más que calor y una pérdida de ATPs con cada turno del ciclo. Tales ciclos se denominan ciclos inútiles y las células tienen controles establecidos para limitar la medida en que ocurren. Dado que los ciclos inútiles pueden, de hecho, producir calor, se utilizan como fuentes de calor en algunos tipos de tejido. El tejido adiposo marrón de los mamíferos utiliza esta estrategia, como se describió anteriormente. Consulta también AQUÍ para más información sobre la generación de calor con un ciclo inútil.
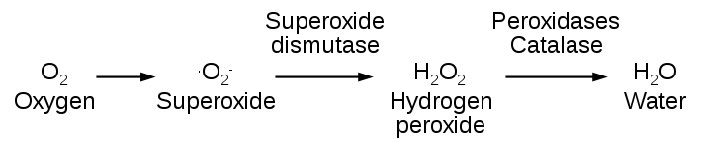
Especies reactivas de oxígeno
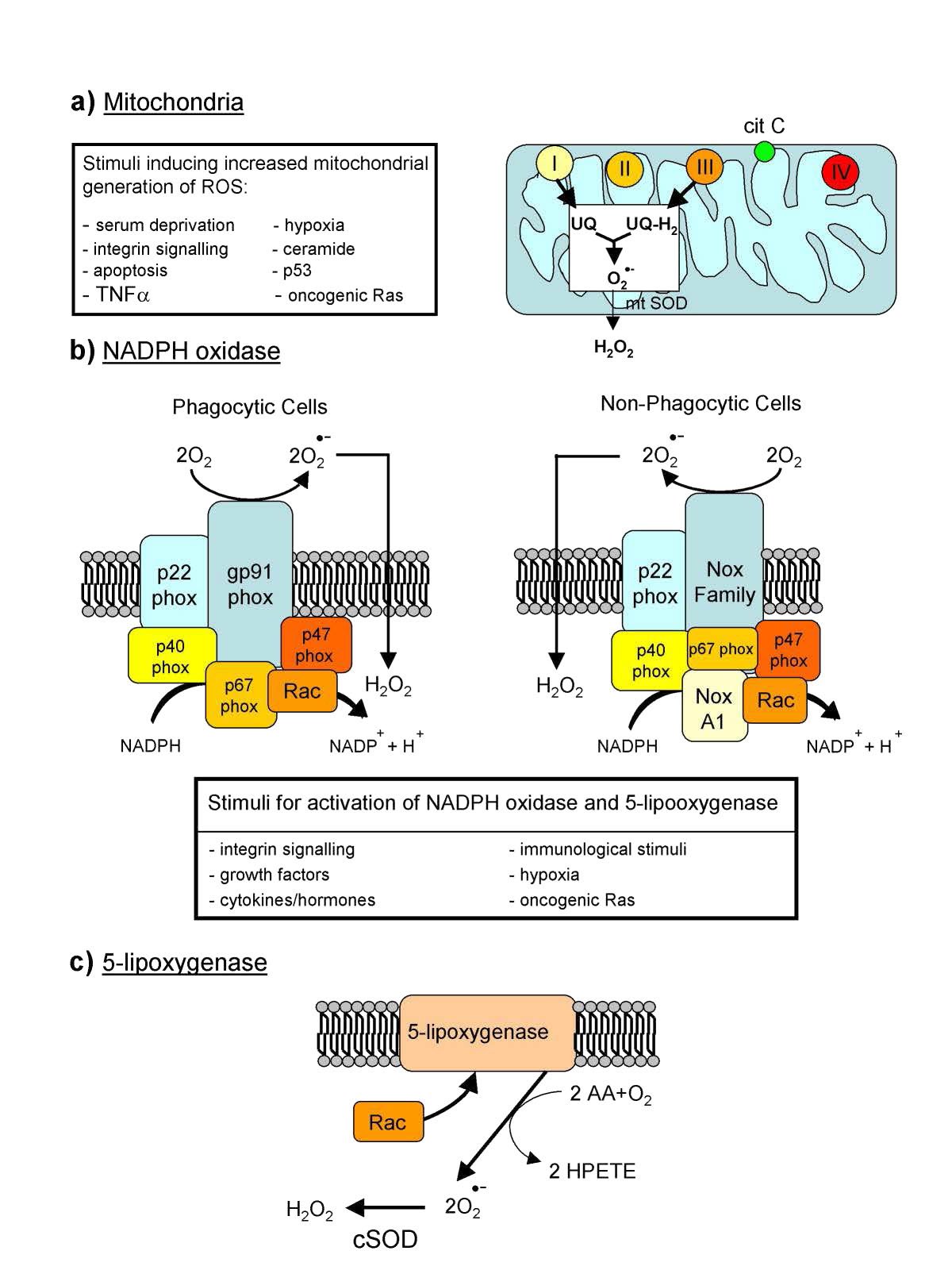
La producción endógena de ROS se dirige hacia la señalización intracelular (H2O2 y óxido nítrico, por ejemplo) y la defensa. Muchas células, por ejemplo, tienen NADPH oxidasa (Figura 5.38) incrustada en la porción exterior de las membranas plasmáticas, en peroxisomas y retículo endoplásmico. Produce superóxidos en la reacción de abajo para matar bacterias.
En el sistema inmune, las células llamadas fagocitos engullen células extrañas y luego usan ROS para matarlas. Las ROS pueden servir como señales para la acción. En el pez cebra, los tejidos dañados han aumentado los niveles de H2O2 y se cree que esto es una señal para que los glóbulos blancos converjan en el sitio. En los peces que carecen de los genes para producir peróxido de hidrógeno, los glóbulos blancos no convergen en el sitio del daño. Las fuentes de peróxido de hidrógeno incluyen los peroxisomas, los cuales lo generan como subproducto de la oxidación de ácidos grasos de cadena larga.
Envejecimiento
Las especies reactivas de oxígeno están en el corazón de la teoría de los radicales libres del envejecimiento, que establece que los organismos envejecen debido a la acumulación de daño de los radicales libres en sus células. En levaduras y Drosophila, hay evidencia de que reducir el daño oxidativo puede aumentar la vida útil. En ratones, el aumento del daño oxidativo disminuye la vida útil, aunque en Caenorhabditis, el bloqueo de la producción de superóxido dismutasa en realidad aumenta la vida útil, por lo que el papel de las ROS en el envejecimiento no está completamente claro.
Sin embargo, está claro que la acumulación de daño mitocondrial es problemática para las células individuales. Las proteínas Bcl-2 en la superficie de las mitocondrias monitorean el daño y si lo detectan, activarán proteínas llamadas Bax para estimular la liberación del citocromo c de la membrana mitocondrial, estimulando la apoptosis (muerte celular programada). Eventualmente la célula muerta será fagocitada.
Una fuente endógena común de superóxido es la cadena de transporte de electrones. El superóxido se puede producir cuando el movimiento de electrones dentro y fuera de la cadena no coincide bien. En estas circunstancias, la CoQ semi-reducida puede donar un electrón al O2 para formar superóxido (O2-). El superóxido puede reaccionar con muchas moléculas, incluido el ADN, donde puede causar daños que conducen a la mutación. Si reacciona con la enzima aconitasa, se puede liberar hierro ferroso (Fe++) que, a su vez, puede reaccionar en la reacción de Fenton para producir otra especie reactiva de oxígeno, el radical hidroxilo (Figura 5.39).
Contrarrestar los efectos de las ROS están las enzimas, como la catalasa, la superóxido dismutasa y los antioxidantes, como el glutatión y las vitaminas C y E.
El glutatión protege contra el daño oxidativo al ser un sustrato para la enzima glutatión peroxidasa. La glutatión peroxidasa cataliza la conversión de peróxido de hidrógeno en agua (página siguiente).
Catalasa
2 H2O2 <=> 2 H2O + O2
La enzima, que emplea cuatro grupos hemo en su catálisis, trabaja de manera extremadamente rápida, convirtiendo hasta 40 mil millones de moléculas de peróxido de hidrógeno en agua y oxígeno por enzima por segundo. Se encuentra abundantemente en los peroxisomas.
Además de la capacidad de la catalasa para descomponer el peróxido de hidrógeno, la enzima también puede usar peróxido de hidrógeno para oxidar una amplia variedad de compuestos orgánicos, incluyendo fenoles, ácido fórmico, formaldehído, acetaldehído y alcoholes, pero con una eficiencia mucho menor.
Salud
La importancia de la catalasa para la salud es incierta. Los ratones deficientes en la enzima parecen sanos y los humanos con niveles bajos de la enzima muestran pocos problemas. Por otro lado, los ratones diseñados para producir mayores niveles de catalasa, en al menos un estudio, vivieron más tiempo. La capacidad de los organismos para vivir con niveles más bajos o sin catalasa puede surgir de otro grupo de enzimas, las peroxiredoxinas, que también actúan sobre el peróxido de hidrógeno y pueden compensar menores cantidades de catalasa. Por último, hay evidencia de que los niveles reducidos de catalasa con el envejecimiento pueden ser responsables del encanecimiento del cabello. Los niveles más altos de H2O2 con catalasa reducida dan como resultado un blanqueamiento de los folículos pilosos.
Superóxido dismutasa
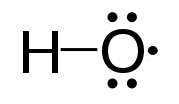
Otra enzima importante para la protección contra especies reactivas de oxígeno es la superóxido dismutasa (SOD), que se encuentra, como la catalasa, en prácticamente todos los organismos que viven en un ambiente de oxígeno. La superóxido dismutasa, también como la catalasa, tiene un valor de Kcat muy alto y, de hecho, tiene el Kcat/Km más alto conocido por cualquier enzima conocida. Cataliza las reacciones en la parte superior de la siguiente columna (superóxidos mostrados en rojo):
La enzima funciona así mediante un mecanismo de ping-pong (doble desplazamiento) (ver AQUÍ), siendo convertida entre dos formas diferentes.
El peróxido de hidrógeno producido en la segunda reacción es fácilmente manejado por la catalasa y además es menos dañino que el superóxido, el cual puede reaccionar con el óxido nítrico (NO) para formar iones peroxinitrito muy tóxicos (Figura 5.43). El peroxinitrito tiene efectos negativos en las células, como se muestra en la Figura 5.45.
Además del cobre, un ion de Zn++ también está unido por la enzima y probablemente juega un papel en la catálisis. También se conocen formas de superóxido dismutasa que usan manganeso, níquel o hierro y se encuentran principalmente en procariotas y protistas, aunque en la mayoría de las mitocondrias se encuentran SOD de manganeso. Las enzimas de cobre/zinc son comunes en eucariotas.
Se encuentran tres formas de superóxido dismutasa en humanos y localizadas en el citoplasma (SOD1 - Figura 5.45), mitocondrias (SOD2 - Figura 5.46) y áreas extracelulares (SOD3 - Figura 5.47). Los ratones que carecen de cualquiera de las tres formas de la enzima son más sensibles a las drogas, como el paraquat. La deficiencia de SOD1 en ratones conduce a carcinoma hepatocelular y pérdida temprana de tejido muscular relacionada con el envejecimiento. Drosophila que carece de SOD2 muere antes del nacimiento y las que carecen de SOD1 envejecen prematuramente.
En humanos, las mutaciones superóxido dismutasa están asociadas con la forma genéticamente ligada de la Esclerosis Lateral Amiotrófica (ELA) y la sobreexpresión del gen está vinculada a trastornos neuronales asociados con el síndrome de Down.
Oxidasas de función mixta
Otras enzimas que catalizan reacciones que involucran oxígeno incluyen las oxidasas de función mixta. Estas enzimas utilizan oxígeno molecular para dos propósitos diferentes en una reacción. La parte de función mixta del nombre se utiliza para indicar reacciones en las que dos sustratos diferentes están siendo oxidados simultáneamente. Las monooxigenasas son ejemplos de oxidasas de función mixta. A continuación se muestra un ejemplo de una reacción de oxidasa de función mixta.
AH + BH2 + O2 <=> AOH + B + H2O
En este caso, la molécula de oxígeno tiene un átomo que sirve como aceptor de electrones y el otro átomo se agrega al AH, creando un alcohol.
Enzimas del citocromo P450
Las enzimas del citocromo P450 (llamadas CYP) son una familia de enzimas oxidasas de función mixta que contienen hemo que se encuentran en todos los dominios de la vida. Se conocen más de 21.000 enzimas CYP. La reacción más característica catalizada por estas enzimas sigue
Las reacciones de monooxigenasas como esta son relativamente raras en la célula debido a su uso de oxígeno molecular. Los CYP requieren un donante de electrones para reacciones como la que se muestra aquí y frecuentemente requieren una proteína para ayudar en la transferencia de electrones para reducir el hierro hemo. Existen seis clases diferentes de enzimas P450 en función de cómo obtienen electrones
1. Bacteriano P450 - electrones de ferredoxina reductasa y ferredoxina
2. Mitocondrial P450 - electrones de la adrenodoxina reductasa y adrenodoxina
3. Cyb5r/Cyb5 - los electrones provienen del citocromo b5
4. FMN/fd - use una FMN reductasa fusionada
5. Microsomal P450 - Los electrones NADPH provienen a través del citocromo P450 reductasa o del citocromo b5 y citocromo b5 reductasa
6. Sistemas solo P450: no requieren potencia reductora externa
Los genes CYP son abundantes en humanos y catalizan miles de reacciones tanto en químicos celulares como extracelulares. Hay 57 genes humanos categorizados en 18 familias diferentes de enzimas. Algunos CYP son específicos para uno o unos pocos sustratos, pero otros pueden actuar sobre muchos sustratos diferentes.
Las enzimas CYP se encuentran en la mayoría de los tejidos del cuerpo y desempeñan funciones importantes en la síntesis de esteroides (colesterol, estrógeno, testoterona, vitamina D, por ejemplo), descomposición de compuestos endógenos (bilirrubina) y en la desintoxicación de compuestos tóxicos incluyendo fármacos. Debido a que actúan sobre muchos fármacos, los cambios en la actividad del CYP pueden producir resultados inesperados y causar problemas con las interacciones medicamentosas.
Los compuestos bioactivos, por ejemplo, en el jugo de pomelo, pueden inhibir la actividad del CYP3A4, lo que lleva a un aumento de las concentraciones circulantes de fármacos sobre los que normalmente habría actuado el CYP3A4. Esta es la razón por la que se aconseja a los pacientes recetados medicamentos que se sabe que son sustratos del CYP3A4 que eviten beber jugo de pomelo mientras están bajo tratamiento. La hierba de San Juan, un tratamiento a base de hierbas, por otro lado, induce la actividad del CYP3A4, pero inhibe CYP1A1, CYP1B1 y CYP2D6. El humo del tabaco induce el CYP1A2 y el berro inhibe el CYP2E1.
Citocromos
Los citocromos son proteínas que contienen hemo que desempeñan un papel importante en el proceso de transporte de electrones en la mitocondria y en la fotosíntesis en el cloroplasto. Existen ya sea como monómeros (citocromo c) o como subunidades dentro de grandes complejos redox (Complejo III y Complejo IV de transporte de electrones. Un átomo de hierro en el centro del grupo hemo juega un papel central en el proceso, volteando entre los estados ferroso (Fe++) y férrico (Fe+++) como resultado del movimiento de electrones a través del mismo.
Existen varios citocromos diferentes. El citocromo c (Figura 5.47) es una proteína soluble poco asociada con la mitocondria. Los citocromos a y a3 se encuentran en el Complejo IV. El complejo III tiene citocromos b y c1 y la plastoquinol-plastocianina reductasa del cloroplasto contiene citocromos b6 y f. Otra clase importante de enzimas que contienen citocromos es el grupo citocromo P450 oxidasa (ver arriba). Obtienen su nombre por el hecho de que absorben luz a 450 nm cuando se reduce su hierro hemo.
Proteínas de hierro y azufre
Las proteínas hierro-azufre contienen grupos de hierro-azufre en una variedad de formatos, incluyendo centros di-, tri- y tetrairon unidos a sulfuro que existen en diferentes estados de oxidación (Figuras 5.48 y 5.49). Los cúmulos juegan una variedad de papeles, pero los más conocidos están en el transporte de electrones donde funcionan en las reacciones redox involucradas en el movimiento de los electrones.
Los Complejos I y el Complejo II contienen múltiples centros Fe-S. Las proteínas de hierro-azufre, sin embargo, tienen muchos otros papeles en las células. La aconitasa utiliza un centro de hierro-azufre en su acción catalítica y la capacidad de la enzima para unirse al hierro le permite funcionar como un barómetro de la concentración de hierro en las células. Los centros de hierro-azufre ayudan a generar radicales en enzimas que utilizan S-Adenosil Metionina (SAM) y pueden servir como fuente de azufre en la síntesis de biotina y ácido lipoico. Algunas proteínas hierro-azufre incluso ayudan a regular la expresión génica.
Ferredoxina
Las ferredoxinas son proteínas que contienen hierro-azufre que realizan transferencia de electrones en una amplia variedad de sistemas y procesos biológicos. Incluyen papeles en la fotosíntesis en cloroplastos. Las ferredoxinas se clasifican estructuralmente por los centros agrupados hierro-azufre que contienen. Los clústeres de Fe2S2 (Figura 5.50) se encuentran en membranas de cloroplastos y pueden donar electrones a glutamato sintasa, nitrato reductasa y sulfito reductasa y sirven como portadores de electrones entre las flavoproteínas reductasa y los sistemas bacterianos de dioxigenasa. La adrenodoxina es una ferredoxina Fe2S2 humana soluble (también llamada ferredoxina 1) que sirve como portador de electrones (al citocromo P450) en sistemas de monooxigenasas mitocondriales. Las ferredoxinas Fe4S4 se subdividen en ferredoxinas de bajo y alto potencial, y estas últimas funcionan en cadenas anaerobias de transporte de electrones.
Ferritina
La ferritina es una proteína intracelular de almacenamiento de hierro que se encuentra en casi todos los organismos vivos, desde bacterias hasta plantas y animales superiores. Es un complejo proteico globular con 24 subunidades y es la principal proteína intracelular de almacenamiento de hierro en eucariotas y procariotas. La ferritina funciona para mantener el hierro en una forma soluble y no tóxica. Su capacidad para almacenar hierro de manera segura y liberarlo de manera controlada le permite actuar como el principal tampón de hierro y solubilizante en las células, evitando que la concentración de hierro libre vaya a alta o caiga demasiado baja. La ferritina se localiza en el citoplasma en la mayoría de los tejidos, pero también se encuentra en el suero actuando como portador de hierro. La ferritina que no contiene hierro se conoce como apoferritina.
Monoaminooxidasas
Las monoaminooxidasas son enzimas que catalizan la desaminación oxidativa de monoaminas, como la serotonina, la epinefrina y la dopamina. La eliminación de la amina con oxígeno da como resultado la producción de un aldehído y amoníaco. Las enzimas se encuentran dentro y fuera del sistema nervioso central.
Existen dos tipos de enzimas monoaminooxidasa: MAO-A y MAO-B. El MAO-A es particularmente importante para oxidar las monoaminas consumidas en la dieta. Tanto MAO-A como MAO-B desempeñan un papel importante en la inactivación de los neurotransmisores monoaminérgicos. Ambas enzimas actúan sobre dopamina, tiramina (Figura 5.50) y triptamina. MAO-A es la enzima principal para metabolizar melatonina, serotonina, norepinefrina y epinefrina, mientras que MAO-B es la enzima primaria para fenetilamina (Figura 5.51) y bencilamina. Se ha reportado que los niveles de MAO-B se reducen considerablemente con el consumo de tabaco.
Las acciones de las monoaminooxidasas afectan así los niveles de neurotransmisores y, en consecuencia, se piensa que desempeñan un papel en los trastornos neurológicos y/o psiquiátricos. Los niveles aberrantes de MAO se han relacionado con numerosos problemas psicológicos, incluyendo depresión, trastorno por déficit de atención (ADD), migrañas, esquizofrenia y abuso de sustancias. Los medicamentos dirigidos a los MAO a veces se usan para tratar la depresión como último recurso, debido a los posibles efectos secundarios. Los niveles excesivos de catecolaminas, como epinefrina, norepinefrina y dopamina, pueden dar lugar a eventos peligrosos de hipertensión.
Teoría del daño al ADN del envejecimiento
La Teoría del Daño al ADN del Envejecimiento se basa en la observación de que, con el tiempo, las células están sujetas a extensos eventos oxidativos. Como ya se señaló, estos brindan oportunidades para la formación de ROS que pueden dañar las moléculas celulares, y se deduce que la acumulación de dicho daño, especialmente al ADN, sería perjudicial para la célula. La acumulación de daños en el ADN podría, por lo tanto, ser responsable de los cambios en la expresión génica que asociamos con el envejecimiento.
Numerosos eventos de daños
La cantidad de daño en el ADN que puede ocurrir es considerable. En ratones, por ejemplo, se estima que cada célula experimenta 40,000 a 150,000 eventos de daño por día. El daño, que ocurre tanto en el ADN nuclear como en el mitocondrial, puede resultar en apoptosis y/o senescencia celular. Los sistemas de reparación del ADN, por supuesto, protegen contra daños al ADN, pero con el tiempo, pueden acumularse daños irreparables.
Daño oxidativo
El daño en el ADN puede ocurrir de varias maneras. La oxidación puede dañar los nucleótidos y alterar sus tendencias de apareamiento de bases. La oxidación de guanina por especies reactivas de oxígeno, por ejemplo, puede producir 8-oxo-guanina (Figuras 5.52 y 5.53). Esta nucleobase oxidada produce comúnmente lesión en el ADN que surge de la acción de especies reactivas de oxígeno como los superóxidos. La 8-oxoguanina es capaz de formar una interacción estable de emparejamiento de bases dentro de un dúplex de ADN con adenina, dando lugar potencialmente a una mutación cuando procede la replicación del ADN. La 8-oxoguanina puede ser reparado si es reconocido en el tiempo por una ADN glicosilasa, que actúa para cortar la base dañada y luego puede ser reemplazada por la apropiada. Los hidrocarburos aromáticos policíclicos del humo del cigarrillo, los gases de escape de diesel o la carne cocida pueden unirse covalentemente al ADN y, si no se reparan, conducir a la mutación. El daño químico al ADN puede resultar en ADN rotos o reticulados.
Enfermedades de la reparación del ADN
La importancia de la reparación del ADN en el proceso de envejecimiento queda clara por enfermedades que afectan la reparación del ADN que conducen al envejecimiento prematuro. Entre ellos se encuentra el síndrome de Werner, para quien la esperanza de vida es de 47 años. Surge como consecuencia de la pérdida de dos enzimas en la reparación por escisión de bases. Las personas que padecen el síndrome de Cockayne tienen una esperanza de vida de 13 años debido a mutaciones que alteran la reparación de la escisión nucleotídica acoplada a la transcripción, que es un sistema importante para la fijación del daño oxidativo.
Además, las expectativas de vida de 13 especies de organismos mamíferos se correlacionan con el nivel de expresión de la proteína inductora de reparación de ADN PARP. Curiosamente, las personas que vivieron más allá de los 100 años tenían un mayor nivel de PARP que las personas más jóvenes de la población.
Antioxidantes
Existe un creciente interés en el tema de los antioxidantes debido a las preocupaciones de salud planteadas por nuestro conocimiento de problemas creados como resultado de la oxidación espontánea de biomoléculas por Especies Reactivas de Oxígeno (ROS), como el superóxido. Los antioxidantes tienen la propiedad química de proteger contra el daño oxidativo al ser fácilmente oxidados ellos mismos, preferentemente a otras biomoléculas.
Biológicamente, las células tienen varias líneas de defensa antioxidante. Incluyen moléculas, como las vitaminas C, A y E, glutatión y enzimas que destruyen las ROS como superóxido dismutasa, catalasa y peroxidasas.

Efectos en la salud
La oxidación por ROS es mutagénica y se ha relacionado con la aterosclerosis. Sin embargo, los estudios aleatorizados de suplementación oral de varias combinaciones de vitaminas no han mostrado ningún efecto protector contra el cáncer y la suplementación de vitamina E y selenio no ha revelado disminución en el riesgo de enfermedad cardiovascular. Además, no se ha encontrado reducción en las tasas de mortalidad como resultado de la suplementación con estos materiales, por lo que los efectos protectores, en su caso, de los antioxidantes sobre las ROS en la salud humana siguen siendo poco conocidos.
Glutatión
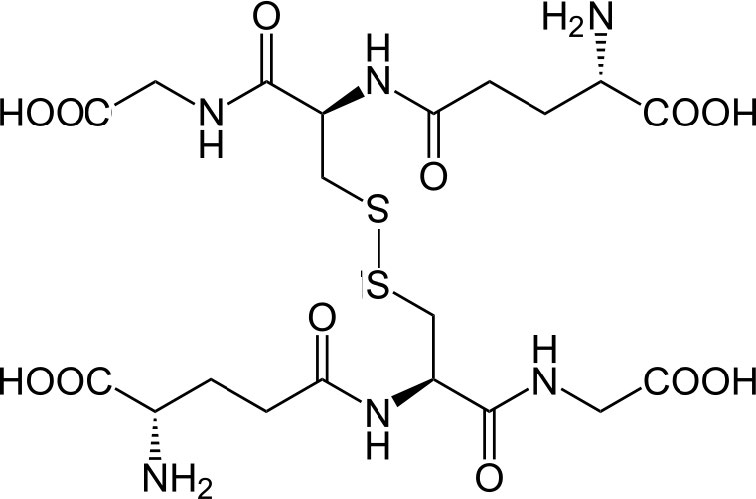
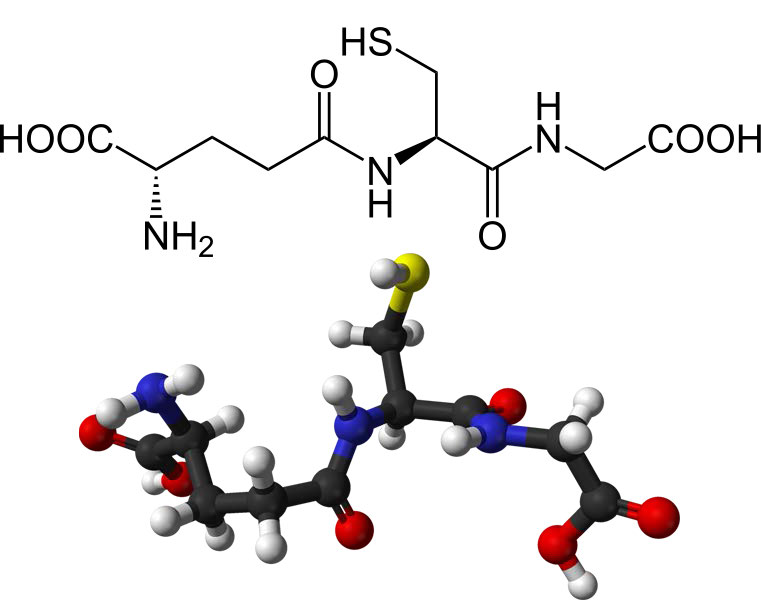
El grupo tiol de la cisteína es un agente reductor que reduce los enlaces disulfuro a sulfhidrilos en proteínas citoplasmáticas. Este, a su vez, es el puente cuando dos glutationes se oxidan y forman un enlace disulfuro entre sí (Figura 5.56). Los dos estados oxidativos del glutatión se abrevian de la siguiente manera: GSH (reducido) y GSSG (oxidado).
Los glutationes unidos por disulfuro se pueden separar por reducción de sus enlaces con glutatión reductasa, utilizando electrones del NADPH para la reducción.
Síntesis no ribosómica
El glutatión no es hecho por los ribosomas. Más bien, dos enzimas catalizan su síntesis. La enzima γ-glutamilcisteína sintetasa cataliza la unión del glutamato a la cisteína y luego la glutatión sintetasa cataliza la formación de enlaces peptídicos entre la cisteína y la glicina. Cada paso requiere energía de ATP.
Esencial para la vida
El glutatión es importante para la vida. Los ratones que carecen de la primera enzima involucrada en su síntesis en el hígado mueren en el primer mes después del nacimiento. En las células sanas, el 90% del glutatión se encuentra en el estado GSH. Los niveles más altos de GSSG corresponden a células que están estresadas oxidativamente.
Además de reducir los enlaces disulfuro en las células, el glutatión también es importante para lo siguiente:
• Neutralización de radicales libres y especies reactivas de oxígeno.
• Mantenimiento de antioxidantes exógenos como las vitaminas C y E en sus formas reducidas.
Regulación del ciclo del óxido nítrico
Resveratrol
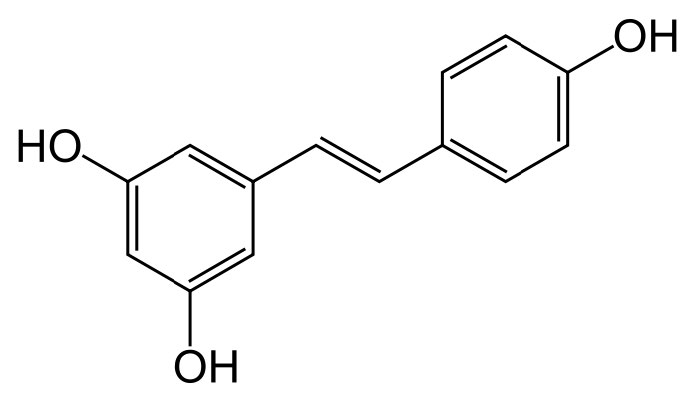
Algunos datos indican que el resveratrol puede mejorar el funcionamiento de las mitocondrias. También actúa como antioxidante y provoca que la concentración de otro antioxidante, el glutatión, aumente. El compuesto parece inducir la expresión de la superóxido dismutasa de manganeso (protege contra especies reactivas de oxígeno) e inhibe varias fosfodiesterasas. Esto provoca un aumento en el AMPc que resulta en aumentos en la oxidación de los ácidos grasos, la formación de mitocondrias, la gluconeogénesis y la descomposición del glucógeno. Se ha afirmado que es la causa de la paradoja francesa en la que se supone que beber vino tinto da protección al sistema cardiovascular. Sin embargo, faltan datos de investigación que respalden la afirmación. Se sabe que el resveratrol activa las proteínas Sirtuina, que desempeñan un papel en la inactivación génica.
Resumen
En resumen, se necesita energía para que las células realicen las funciones que deben realizar para mantenerse con vida. En su nivel más básico, esto significa librar una batalla continua con la entropía, pero no es la única necesidad de energía que tienen las células.
Referencias
1. Winge, D.R., Mol Cell Biol. 2012 Jul; 32 (14): 2647—2652. doi: 10.1128/MCB.00573-12
Energía: Transporte de electrones y fosforilación oxidativa
429
Conferencias de YouTube
por Kevin
AQUÍ Y AQUÍ
Conferencias de YouTube
por Kevin
AQUÍ Y AQUÍ
430
Figura 5.14 - Visión general del transporte de electrones (abajo a la izquierda y arriba a la derecha) y la fosforilación oxidativa (arriba a la izquierda - caja amarilla) en la mitocondria
431
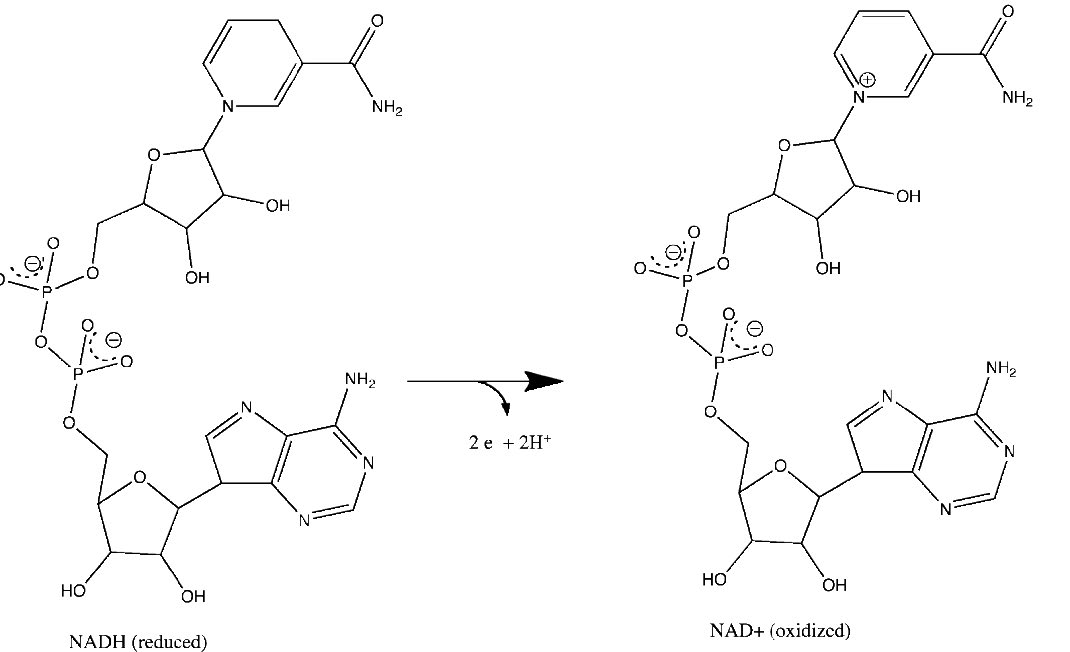
Figura 5.15 - Pérdida de electrones por NADH para formar NAD+. Las reacciones relevantes ocurren en el anillo superior de la molécula.
432
Figura 5.16 - Flujo de electrones del NADH al sistema de transporte de electrones. La entrada es a través del complejo I
Imagen de Aleia Kim
Figura 5.17 - Flujo de electrones desde FADH2 hacia la cadena de transporte de electrones. La entrada es a través del complejo II.
Imagen de Aleia Kim
Aprendizaje Interactivo
Módulo
AQUÍ
433
Figura 5.18 - Complejo I incrustado en la membrana mitocondrial interna. La matriz mitocondrial en la parte superior
Wikipedia
434
Figura 5.19 - Complejo II embebido en la membrana mitocondrial interna. Matrix está arriba.
Wikipedia
Conferencias de YouTube
por Kevin
AQUÍ Y AQUÍ
435
Figura 5.20 - Movimiento de electrones a través del complejo I del NADH a la coenzima Q. La matriz mitocondrial está en la parte inferior
Imagen de Aleia Kim
Figura 5.21 - Movimiento de electrones desde succinato a través del complejo II (A->B->C->D->Q). Matriz mitocondrial en la parte inferior.
Imagen de Aleia Kim
436
Figura 5.22 - Complejo II en membrana mitocondrial interna que muestra flujo de electrones. Matrix está arriba.
Wikipedia
Figura 5.23 - Coenzima Q
437
Película 5.2 - El ciclo Q
Wikipedia
Figura 5.24 - La imagen del ciclo Q por Aleia Kim
Figura 5.24 - Complejo III
Wikipedia
438
Conferencias de YouTube
por Kevin
AQUÍ Y AQUÍ
Figura 5.25 - El ciclo Q. Matrix está abajo.
Imagen de Aleia Kim
439
Figura 5.26 - Movimiento de electrones y protones a través del complejo IV. Matrix está abajo
Imagen de Aleia Kim
Figura 5.25 - Citocromo c con grupo hemo unido
Wikipedia
440
Figura 5.27 - Anatomía mitocondrial. Los complejos de transporte de electrones y la ATP sintasa están incrustados en la membrana mitocondrial interna
Imagen de Aleia Kim
441
Figura 5.28 - ATP sintasa. Los protones pasan desde el espacio intermembrana (arriba) a través del complejo y salen en la matriz (abajo).
Imagen de Aleia Kim
Aprendizaje Interactivo
Módulo
AQUÍ
442
Película 5.3 - ATP Sintasa - ADP + Pi (rosa) y ATP (rojo). La vista es final desde el lado citoplásmico viendo las subunidades β
Película 5.3 - ATP Sintasa - ADP + Pi (rosa) y ATP (rojo). La vista es final desde el lado citoplásmico viendo las subunidades β
443
Figura 5.29 - Características estructurales importantes de la ATP sintasa
Imagen de Aleia Kim
444
Figura 5.30 - Estructuras Sueltas (L), Apretadas (T) y Abiertas (O) de la cabeza F1 de ATP sintasa. El cambio de estructura ocurre por rotación de la γ-proteína (púrpura) en el centro como resultado del movimiento de protones. Las unidades individuales α y β no giran
Imagen de Aleia Kim
445
Figura 5.31 - Visión general de la respiración en células eucariotas
Wikipedia
Conferencias de YouTube
por Kevin
AQUÍ Y AQUÍ
446
Resto
ATP Alto/ADP Bajo
Fosforilación Oxidativa
Transporte de electrones bajo
Bajo consumo de oxígeno
NADH Alto/NAD+ Bajo
Ciclo de ácido cítrico lento
Ejercicio
ATP Bajo/ADP Alto
Alta fosforilación oxidativa
Alto Transporte de Electrones
Uso de Oxígeno Alto
NADH Bajo/NAD+ Alto
Ciclo de ácido cítrico rápido
Aprendizaje Interactivo
Módulo
AQUÍ
447
Figura 5.32 - Tres inhibidores del transporte de electrones
Imagen de Aleia Kim
448
Figura 5.33 - Oligomicina A - Un inhibidor de ATP sintasa
Figura 5.34 - 2,4 DNP - un desacoplador de control respiratorio
449
En Celdas Con Acoplamiento Estrecho
El uso de O2 depende del metabolismo Los niveles de
NAD+ varían con el ejercicio Gradiente de
protones alto sin ejercicio El
catabolismo depende de las necesidades energéticas
ETS se ejecuta cuando OxPhos corre y viceversa
En celdas que están desacopladas
O2 usa alto
NAD+ Niveles altos
Poco o ningún gradiente de protones
Catabolismo alto
OxPhos no corre, pero ETS corre rápidamente
Conferencias de YouTube
por Kevin
AQUÍ Y AQUÍ
450
451
Figura 5.35 - La oxidasa alternativa (AOX) de hongos, plantas y protozoos evita parte del transporte de electrones tomando electrones de CoQ y pasándolos al oxígeno.
452
Figura 5.36 - Estructura de un radical libre de oxígeno
Wikipedia
NADPH + 2O2
NADP+ + 2O2− + H+
Figura 5.37 - Tres fuentes de especies reactivas de oxígeno (ROS) en células
Wikipedia
453
454
Conferencias de YouTube
por Kevin
AQUÍ Y AQUÍ
Figura 5.38 Un radical hidroxilo
Wikipedia
455
Glutatión reducido (GSH) + H2O2
Glutatión oxidado (GSSG) + H2O
Figura 5.40 - Desintoxicación de especies reactivas de oxígeno
Figura 5.39 - Catalasa
456
1. O2- + Enzima Cu++
O2 + Enzima Cu+
2. O2- + Enzima-Cu+ + 2H+
H2O2 + Enzima-Cu++
Figura 5.41 - SOD2 de humanos
Figura 5.42 3 - Ion de peroxinitrito
Figura 5.44 - SOD1 de humanos
Wikipedia
Figura 5.45 - SOD3 de humanos
457
Figura 5.43 - Los efectos del peroxinitrito en las células conducen a necrosis o apoptosis
Wikipedia
458
RH + O2 + NADPH + H+
ROH + H2O + NADP+
459
Figura 5.46 - Citocromo c con su grupo hemo
460
Conferencias de YouTube
por Kevin
AQUÍ Y AQUÍ
Figura 5.47 - Clúster Fe2S2
Figura 5.48 - Reacciones redox para agrupaciones de Fe4S4
461
Figura 5.49 - Tiramina
Figura 5.50 - Fenetilamina
462
Figura 5.51 - Guanina y 8-oxo-guanina
Figura 5.52 - Par de bases adenina-8-oxo-guanina. dR = desoxirribosa
463
Figura 5.53 - Buenas fuentes de antioxidantes
464
Figura 5.55 - Glutationes oxidados (GSSG) unidos por un enlace disulfuro
Wikipedia
Figura 5.54 - Estructura del glutatión reducido (GSH)
465
Figura 5.56 - Resveratrol
Conferencias de YouTube
por Kevin
AQUÍ Y AQUÍ
466
Las imágenes gráficas de este libro fueron producto del trabajo de varios estudiantes talentosos. Los enlaces a sus páginas web están a continuación
Haga clic AQUÍ para
Martha Baker
Página Web
Haga clic AQUÍ para
Pehr Jacobson
Página Web
Haga clic AQUÍ para
Aleia Kim
Página Web
Haga clic AQUÍ para
Penélope Irving
Página Web
Conjunto de problemas relacionados con esta sección AQUÍ
Resumen punto por punto de esta sección AQUÍ
Para obtener un certificado para dominar esta sección del libro, haga clic AQUÍ
Cursos gratuitos de Kevin Ahern en iTunes U - Básico/Escuela de Medicina/Avanzado
Bioquímica Gratis & Fácil (nuestro otro libro) AQUÍ/Página de Facebook
Guía de Kevin e Indira para ingresar a la escuela de medicina - iTunes U Course/Book
Para ver los cursos de ecampus OSU de Kevin Ahern - BB 350/BB 450/BB 451
Para inscribirse en los cursos OSU ecampus de Kevin Ahern - BB 350/BB 450/BB 451
Bioquímica Gratis Para Todos Página de Facebook (por favor, dale Me Gusta)
Página web de Kevin Ahern/Página de Facebook/Página web de Taralyn Tan
Descargas gratis de Kevin Ahern AQUÍ
Programa de Bioquímica/Biofísica de OSU AQUÍ
Facultad de Ciencias de OSU AQUÍ
Universidad Estatal de Oregón AQUÍ
Correo electrónico Kevin Ahern/Indira Rajagopal/Taralyn Tan
Soy un poco mitocondria
Quien te da energia
Yo uso mi gradiente de protones
Para hacer los ATP
Es un poco mitocondria
Quien nos da energia
Utiliza gradientes de protones
Para hacer algunos ATP
Fluyen electrones a través del Complejo II
A policía de tránsito Co-Q
Siempre que lleguen allí en
Un FADH-Dos
electrones fluyen a través del Complejo II Al
policía de tránsito Co-Q
Siempre que llegan ahí en
An Fadh-dos
Estrechamente acoplado es mi estado
A menos que me salga un agujero
Creado en mi membrana por
Algunos di-ni-tro-phe-nol
Si bien acoplado es su estado
A menos que consiga un agujero
Creado en su membrana por
Algunos di-ni-tro-fenol
Tanto la rotenona como el cianuro
Detener mi flujo de electrones
Y detener el cálculo de
Mi relación “P” a “O”
Grabación por Tim Karplus
Letras de Kevin Ahern
Recording por Tim Karplus Letras por Kevin Ahern
Soy una pequeña mitocondria
A la melodía de “Soy Leñador”
Melodías metabólicas Sitio web AQUÍ
En las vías catabólicas que emplean nuestras células Las
oxidaciones ayudan a crear el ATP
mientras bajan la energía libre de Gibbs
Gracias a la entalpía
Si un sustrato se convierte de un alcohol
A un aldehído o cetona está claro
Esos electrones no desaparecen Simplemente
se reordenan — muy extraño
N-A-D está en mis oídos y en mis ojos
Ayudar a mol-e-cules obtener oxidado
Haciendo N-A-D-H entonces
Y esto último es un problema anaeróbicamente
'Cuz acumulaciones de ella
odian los músculos Responden usando piruvato
Para producir lactato
Catalizar es esencial para que las células vivan
Así que las enzimas agarran sus sustratos ansiosamente
Si se unen con alta afinidad
Low Km ya ves, así como yo
N-A-D está en mis oídos y en mis ojos
Help-ing mol-e-cules se oxidan
Haciendo N-A-D-H entonces
N-A-D
Al ritmo de “Penny Lane”
Melodías metabólicas Sitio web AQUÍ
Grabado por Tim Karplus
Letras por Kevin Ahern
Grabada por Tim Karplus Letras por Kevin Ahern
Cuando los electrones del oxígeno están todos en el estado equilibrado
Hay doce de ellos para oh-dos. La molécula es genial
Pero a veces ocurren problemas en la ruta al complejo IV
Hacer especies reactivas que la célula no puede ignorar
Oh, la superóxido dismutasa es súper catalítica
Impedir que las células se vuelvan muy peroxinitríticas
Más rápido que un radical, sus acciones son fabulosas
La superóxido dismutasa es súper catalítica
Enzima, enzima en el interior
Bloqueando todos los malos óxidos
La principal ventaja de la enzima es que no tiene que esperar
Al unir superóxido en un estado de transición cercana
Se convierte en un oxígeno en el mecanismo uno
Produciendo “h dos oh dos” cuando el ciclo está todo terminado
Oh superóxido dismutasa eres más rápido que todos ellos
Tienes la proporción más alta de kcat sobre KM
Esto significa que el superóxido no puede causar demasiado desorden
La superóxido dismutasa es más rápida que todas ellas
Superóxido dismutasa
Detener las formas de superóxido
La enzima es como una pelota de ping-pong que mecanicista
Rebotan entre dos estados de cobre, más uno y dos ves
Entonces S-O-D se comporta igual que un antioxidante
Dando tanta protección como una célula podría desear
Oh superóxido dismutasa, la célula está enamorada de ti
Porque dejas que el transporte de electrones haga lo que debe hacer
Sin acumulación de un radical oh dos
Superóxido dismutasa - por eso te ama una célula
Superóxido dismutasa
A la melodía de “Supercalifragilistiexpialidocious”
Melodías metabólicas Sitio web AQUÍ
Letras por Kevin Ahern
Aún no hay grabación para esta canción