
16.3: Competencia de recursos
- Page ID
- 53267

La competencia entre dos especies ocurre cuando los términos de interacción\(s_{1,2}\) y\(s_{2,1}\) en la Ecuación 8.1 son negativos. Este enfoque bastante abstracto nos lleva a una visión amplia, pero para otros tipos de percepciones pasemos a una visión más mecanicista. En lugar de coeficientes abstractos que representan la inhibición entre especies, consideremos los recursos que necesitan para prosperar y sobrevivir. Las especies no interactuarán directamente, ni siquiera necesitan entrar en contacto, sino que se influirán mutuamente a través del uso de un recurso común, que ambos necesitan para el mantenimiento y crecimiento de sus poblaciones.
La competencia de recursos es una de las partes más antiguas de la teoría ecológica, introducida a finales de la década de 1920 por el matemático Vivo Volterra. Empezaremos por donde empezó, considerando lo que se ha denominado “recursos abióticos”.
Las especies requieren luz solar, espacio, nitrógeno, fósforo y otros recursos en diversas cantidades. Si un recurso es demasiado raro, las poblaciones no pueden crecer, y de hecho disminuirán. En la Figura\(\PageIndex{1}\) esto se muestra en la región a la izquierda de la flecha marcada A, en la que la tasa de crecimiento individual\(1/N\,dN/dt\) es negativa.
A mayores niveles de recursos la tasa de crecimiento aumenta y, en el punto A, la población apenas puede mantenerse. Aquí la tasa de crecimiento individual\(1/N\,dN/dt\) es cero. A este nivel de recursos se le llama\(R^{\ast}\), se pronuncia “son estrellas”. A mayores niveles de recursos, arriba\(R^{\ast}\), la población crece porque\(1/N\,dN/dt\) se vuelve positiva.
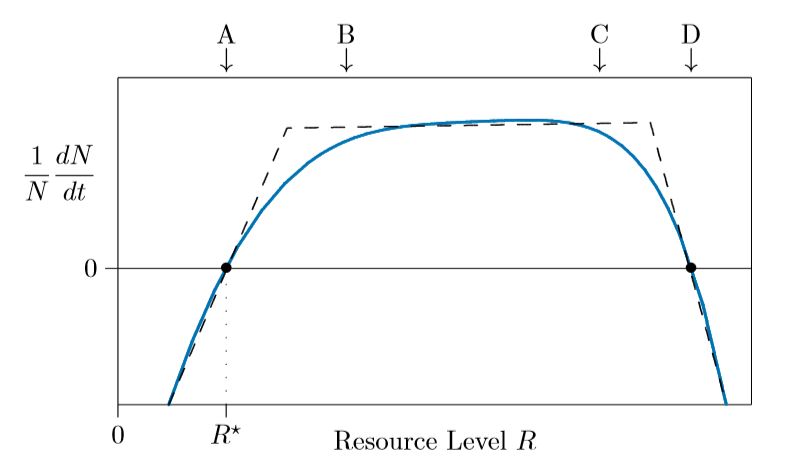
Una vez que los recursos son abundantes, aproximadamente por encima de B en la figura, las necesidades se sacian y la adición de más recursos no hace ninguna gran diferencia. El crecimiento poblacional se mantiene aproximadamente igual entre las marcas B y C.
En niveles muy altos, demasiados recursos pueden dañar realmente a la población. Demasiada luz solar puede quemar las hojas, por ejemplo, mientras que demasiado nitrógeno puede dañar las raíces. En este punto, por encima de C en la Figura\(\PageIndex{1}\), la tasa de crecimiento comienza a caer. Por D la especie nuevamente apenas puede sostenerse, y a la derecha de D la especie es asesinada por una sobreabundancia de recursos.
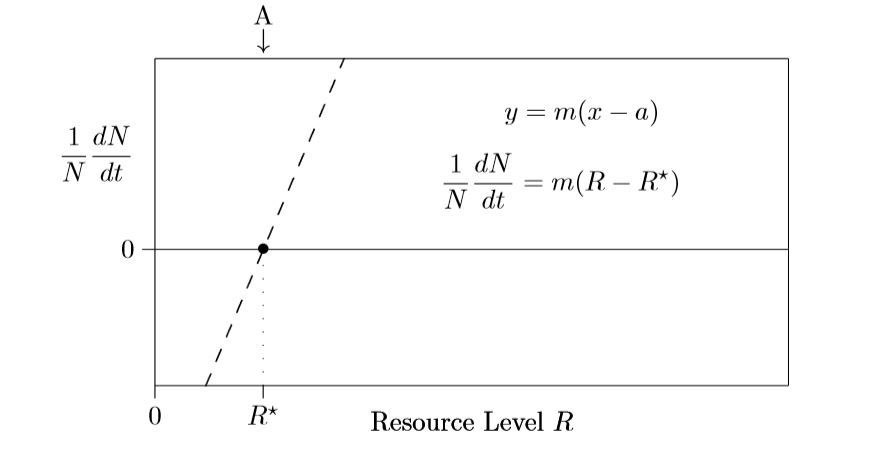
Estos altos niveles de recursos, sin embargo, no suelen observarse, ya que las especies reducen las concentraciones de recursos utilizándolos hacia arriba. A menos que se estén modelizando entornos extremos, solo se\(\PageIndex{1}\) necesita modelar la pieza lineal discontinua izquierda de la Figura, como se muestra en la Figura\(\PageIndex{2}\).
En este punto, es útil revisar diversas formas para la ecuación de una línea recta. La pendiente habitual, la forma de intercepción\(y\,=\,mx+b\), que es una forma de intercepción y, no es tan útil aquí. Es la forma de intercepción x,\(y\,=\,m(x−a)\), la que entra en juego para escribir un modelo mecanicista de recursos para el crecimiento poblacional de una sola especie.
|
1. Pendiente: forma de intercepción:\(y\,=\,mx\,+\,b\) (pendiente m, intercepción y b) 2. Intercept—Intercept form:\(y\,=\,b(1\,−\,x/a)\) (intercepción x a, intercepción y b) (m = −b/a) 3. Pendiente: forma de intercepción X:\(y\,=\,m\,(x − a)\) (pendiente m, intercepción x a) (b = −m/a) |
El punto de crecimiento cero,\(R^{\ast}\), es importante en la teoría de la competencia de recursos. Es la cantidad de recurso que apenas apenas sostiene a la especie. Si el nivel de recursos es menor que\(R^{\ast}\), la especie se extingue; si es mayor, la especie crece y se expande. Por lo tanto, se espera que el nivel de recursos en el medio ambiente sea igual o próximo al\(R^{\ast}\) valor de la especie dominante. Si está por encima de ese nivel, la población crece, los nuevos individuos utilizan más recursos y, en consecuencia, el nivel de recursos se reduce hasta que el crecimiento se detiene.
\(R^{\ast}\)se puede medir en el invernadero o en el campo. En el invernadero, por ejemplo, podrías organizar las plantas en 20 macetas y darles diferentes cantidades de fertilizante nitrogenado en suelo estéril y libre de nutrientes. En ausencia total de fertilizante, las plantas morirán. Con mayores cantidades de fertilizante las plantas serán lujosas, y cuando haya demasiado fertilizante, el recurso se volverá tóxico, lo que volverá a provocar la muerte en las plantas. Así se puede medir la curva de Figura con\(\PageIndex{1}\) bastante facilidad y encontrar el punto de la izquierda donde las plantas simplemente sobreviven. Este es su\(R^{\ast}\).
También puede medir este valor para diferentes especies de forma independiente, y a partir de los resultados estimar cómo les va a ir a las plantas viviendo juntas. Para comenzar, supongamos que un recurso es el más limitante. Representar la cantidad de ese recurso limitante disponible en el entorno en el momento\(t\) por el símbolo\(R(t)\) o, para taquigrafía, simplemente\(R\). La cantidad superior a las necesidades mínimas es\(R−R^{\ast}\), y esa cantidad de exceso determinará la tasa de crecimiento. Un pequeño exceso significará un crecimiento lento, pero un exceso mayor puede soportar un crecimiento más rápido. Entonces la ecuación en la Figura\(\PageIndex{2}\) muestra la tasa de crecimiento individual,\(1/N\,dN/dt\), siendo proporcional a la cantidad de recursos que existe en exceso.
Como antes,\(N\) mide el tamaño de la población en el momento\(t\), en número de individuos, biomasa total, o cualquier unidad que sea relevante para la especie en estudio. \(R^{\ast}\)es la menor cantidad de recursos que puede soportar una población viable, y m dice cómo la tasa de crecimiento individual,\((1/N)dN/dt\), depende de la cantidad de recursos disponibles en exceso de las necesidades mínimas.
Ahora digamos que\(R_{max}\) es la cantidad máxima de recurso disponible en el medio ambiente, en ausencia de algún organismo, y u es la cantidad de recurso que utiliza cada organismo vivo en la población. Entonces\(uN\) es la cantidad de recursos amarrados en la población en el momento\(t\). \(R_{max}\,−\,uN\,=\,R\)es la cantidad de recursos no utilizados por la población. Esta es la base de una teoría de recursos que asume que los recursos se liberan inmediatamente después de la muerte de un organismo, y tiene muchas de las propiedades importantes de modelos de recursos más complejos.
Comience con las declaraciones anteriores en forma algebraica,
\[\frac{1}{N}\frac{dN}{dt}\,=\,m(R-R^{\ast})\,\,\,and\,\,\,R\,=\,R_{max}\,-uN\]
luego sustituya la ecuación de la derecha de arriba por la de la izquierda. Esto da
\(\frac{1}{N}\frac{dN}{dt}\,=\,m\,(R_{max}\,-\,uN\,-\,R^{\ast})\)
Multiplicar a través de la derecha por\(m\) y reorganizar términos da
\[\frac{1}{N}\frac{dN}{dt}\,=\,m(R_{max}\,-\,R^{\ast})\,-\,umN\]
Observe que el primer término a la derecha es una constante y el segundo término es un tiempo constante N. ¿Esto le resulta familiar? Esto es solo crecimiento poblacional regulado por densidad disfrazada, ¡el\(r\,+\,sN\) modelo otra vez!
\(\frac{1}{N}\frac{dN}{dt}\,=\,r+sN,\,\,\,r\,=\,m(R_{max}\,-\,R^{/ast}),\,\,\,s\,=\,-um\)
Recordemos que esto también sucedió para el\(I\) modelo epidemiológico. Y surgirá ocurrir de nuevo en modelos futuros.