En esta conferencia aprendimos cómo las marcas de cromatina pueden ser utilizadas para inferir estados biológicamente relevantes. El análisis en [7] presenta un método sofisticado para aplicar técnicas previamente aprendidas como los HMM a un problema complejo. La conferencia también presentó la poderosa transformación Burrows-Wheeler que ha permitido el mapeo de lectura eciente.
Bibliografía
[1] Langmead B, Trapnell C, Pop M y Salzberg S. Ultrafast, alineamiento memory-ecient de secuencias cortas de ADN con el genoma humano. Biología del Genoma, 10 (3), 2009.
[2] Roadmap Epigenomics Consortium, Kundaje A, Meuleman W, et al. Análisis integrador de 111 epigenomas humanos de referencia. Naturaleza, 518 (7539) :317—330, 2015.
[3] El Consorcio del Proyecto ENCODE. Una enciclopedia integrada de elementos de ADN en el genoma humano. Naturaleza, 489 (7414) :57—74, 2012.
[4] Escuché E y Martienssen RA. Herencia epigenética transgeneracional: Mitos y mecanismos. Cell, 157 (1) :95—109, 2014.
[5] Mardis ER. Chip-seq: bienvenido a la nueva frontera. Nature Methods, 4 (8) :614—614, 2007.
[6] Herz H-M, Hu D y Shilatifard A. Mal funcionamiento del potenciador en cáncer. Molecular Cell, 53 (6) :859—866, 2014.
[7] Ernst J y Kellis M. Descubrimiento y caracterización de estados de cromatina para la anotación sistemática del genoma humano. Nature Biotechnology, 28:817 —825, 2010.
[8] Ernst J, Kheradpour P, Mikkelsen TS, et al. Mapeo y análisis de la dinámica del estado de la cromatina en nueve tipos celulares humanos. Naturaleza, 473 (7345) :43—49, 2011.
[9] Mousavi K, Zare H, dell'Orso S, Grontved L, et al. ERnas promueven la transcripción estableciendo accesibilidad a la cromatina en loci genómicos definidos. Molecular Cell, 51 (5) :606—17, 2013.
[10] Qunhua Li, James B. Brown, Haiyan Huang y Peter J. Bickel. Medición de la reproducibilidad de experimentos de alto rendimiento. Los anales de la estadística aplicada, 5 (3) :1752—1779, 2011.
[11] Li Y y Tollefsbol TO. Detección de metilación del ADN: Análisis de secuenciación genómica con bisulfito. Métodos Biología Molecular, 791:11 —21, 2011.
Cortesía de Institutos Nacionales de Salud. Imagen en el dominio público (izquierda).
Cortesía de Elsevier, Inc. Usado con permiso. Fuente: Herz, Hans-Martin, Deqing Hu, et al. “Mal Mal Mejorador en Cáncer”. Molecular Cell 53, núm. 6 (2014): 859-66 (derecha).
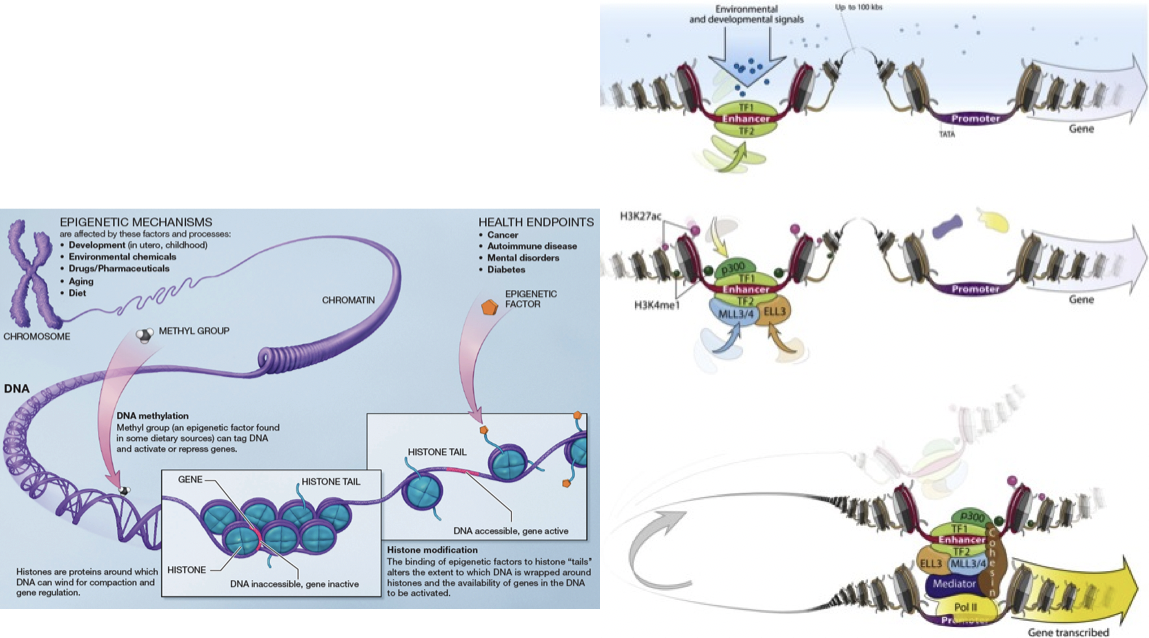
Figura 19.1: A. Existe una amplia diversidad de modificaciones en el epigenoma. Algunas regiones del ADN se enrollan de forma compacta alrededor de las histonas, haciendo que el ADN sea inaccesible y los genes inactivos. Otras regiones tienen ADN más accesible y, por lo tanto, genes activos. Los factores epigenéticos pueden unirse a las colas de estas histonas para modificar estas propiedades. B. Las modificaciones de histonas proporcionan información sobre qué tipos de proteínas están unidas al ADN y cuál es la función de la región. En este ejemplo, las modificaciones de histonas permiten que una región potenciadora (potencialmente a más de 100 bases de kilo de distancia) interactúe con la región promotora. [6]
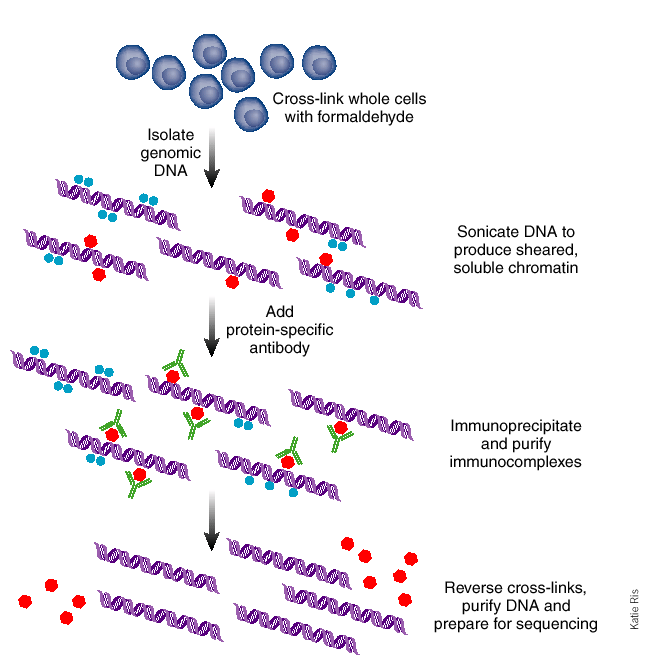
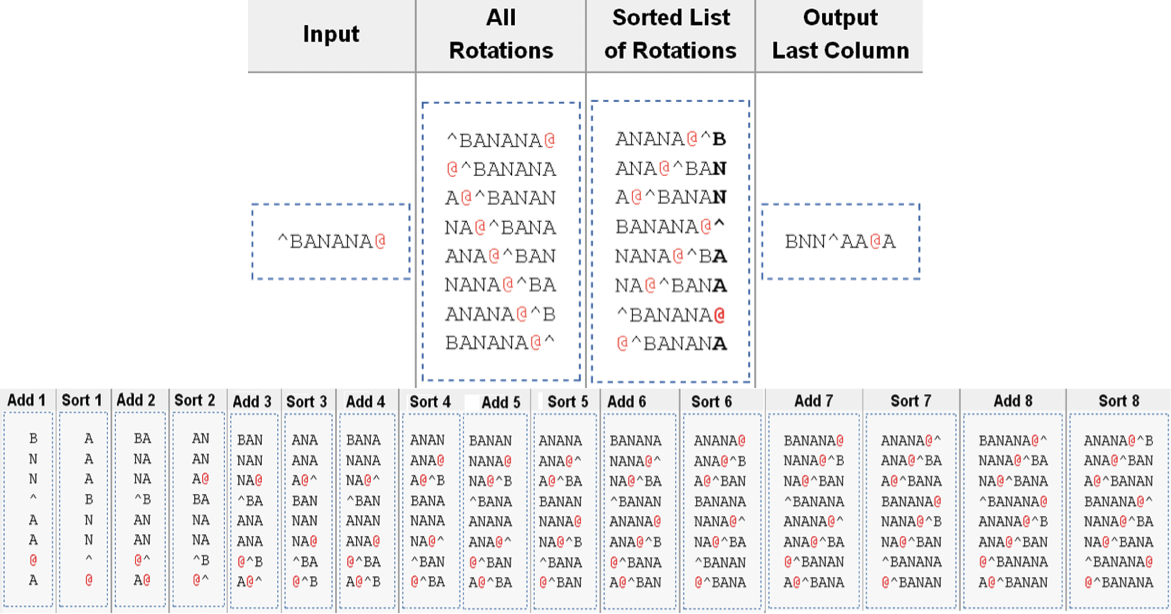
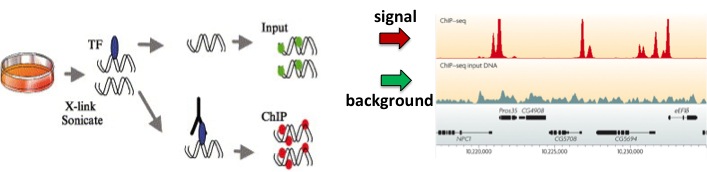
Figura 19.2: El método de inmunoprecipitación de cromatina [5]. Los pasos en esta figura corresponden a los seis pasos del procedimiento.Figura 19.3: (Arriba) En la transformación hacia adelante Madrows-Wheeler se generan y clasifican las rotaciones. La última columna de la lista ordenada (en negrilla) consiste en la cadena transformada. (Abajo) En la transformación inversa de Burrows-Wheeler se ordena la cadena transformada, y se generan dos columnas: una que consiste en la cadena original y la otra compuesta por la ordenada. Éstas forman de manera eficaz dos columnas a partir de las rotaciones en la transformación hacia adelante. Este proceso se repite hasta que se generan las rotaciones completas.Figura 19.4: Para usar ADN de entrada como control, se puede ejecutar el experimento ChIP como normal mientras simultáneamente se ejecuta el mismo experimento (con el mismo ADN) sin un anticuerpo. Esto genera una señal de fondo para la cual podemos corregir.
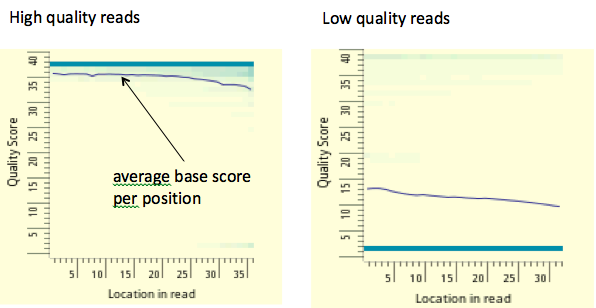
Figura 19.5: En la figura anterior cada columna se encuentra un histograma codificado por colores que codifica la fracción de todas las lecturas mapeadas que tienen puntuación base Q (eje y) en cada posición (eje x). Un puntaje promedio bajo por base implica una mayor probabilidad de errores de mapeo. Normalmente rechazamos lecturas cuya puntuación promedio Q es menor a 10.fuente desconocida. Todos los derechos reservados. Este contenido está excluido de nuestra licencia Creative Commons. Para obtener más información, consulte http://ocw.mit.edu/help/faq-fair-use/.
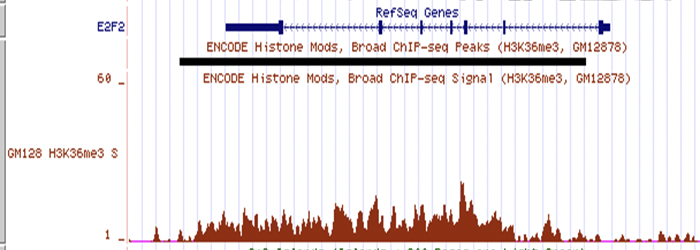
Figura 19.6: Una pista de señal de muestra. Aquí, la señal roja se deriva del número de lecturas que mapearon al genoma en cada posición para un experimento ChIP-seq con la diana H3K36me3. La señal da un nivel de enriquecimiento de la marca
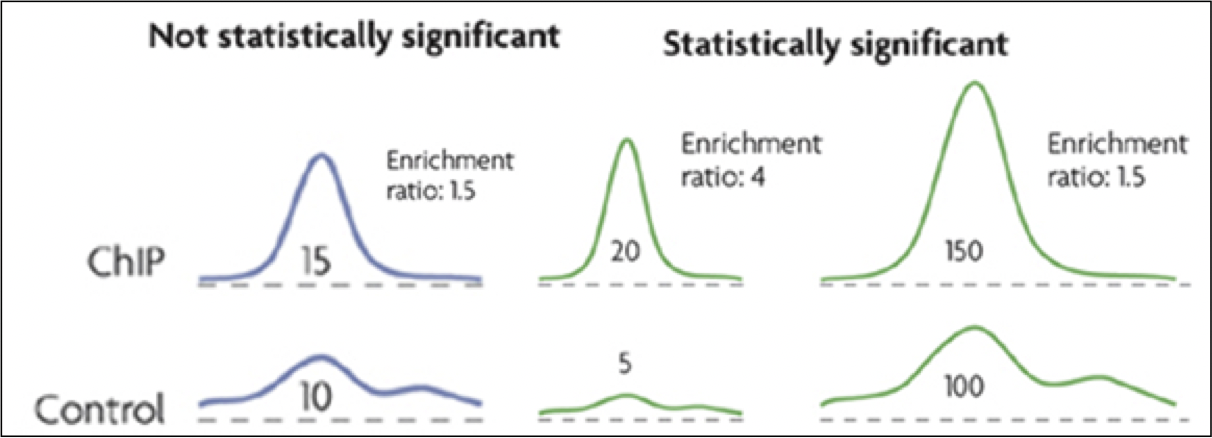
Figura 19.7: Pistas de señal de muestra tanto para el experimento verdadero como para el fondo (control). Se considera que las regiones tienen enriquecimiento estadísticamente significativo cuando los valores verdaderos de la señal del experimento están muy por encima de los valores de la señal de fondo.Cortesía de Macmillan Publishers Limited. Usado con permiso. Fuente: Ernst, Jason y Manolis Kellis. “Descubrimiento y Caracterización de los Estados Cromatinos para la Anotación Sistemática del Genoma Humano”. Nature Biotechnology 28, núm. 8 (2010): 817-25.
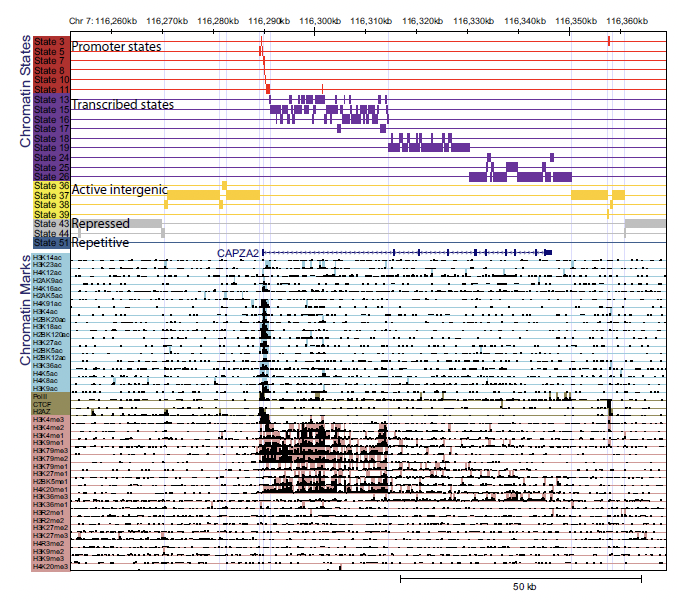
Figura 19.8: Ejemplo de los datos y la anotación del modelo HMM. La sección inferior muestra el número bruto de lecturas mapeadas al genoma. La sección superior muestra la anotación del modelo HMM.
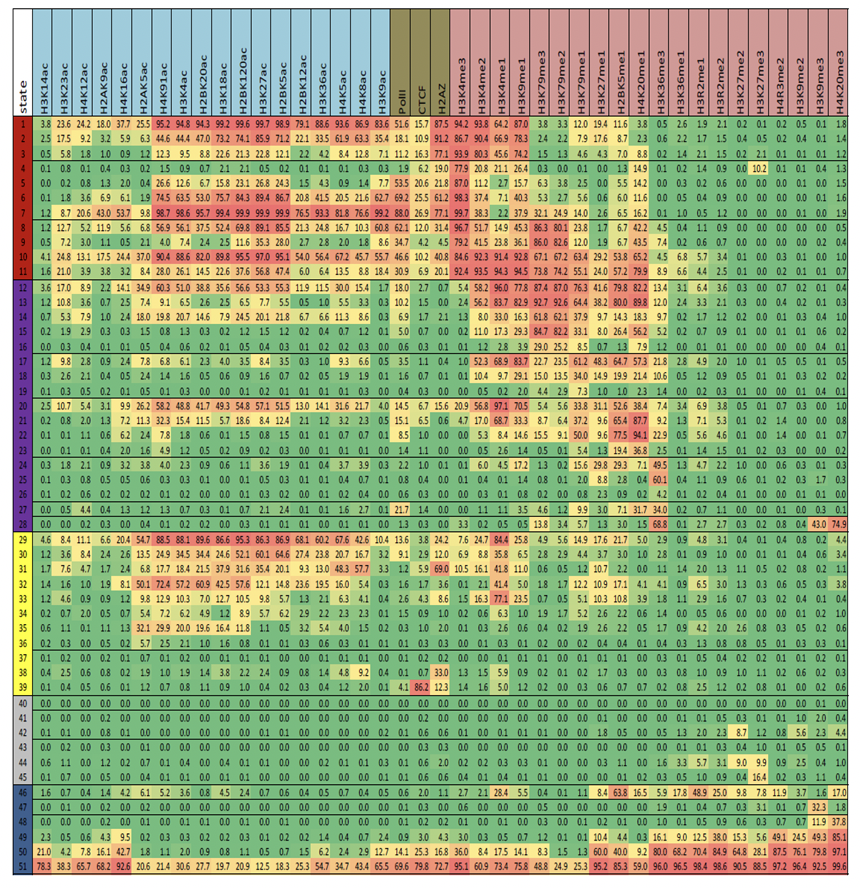
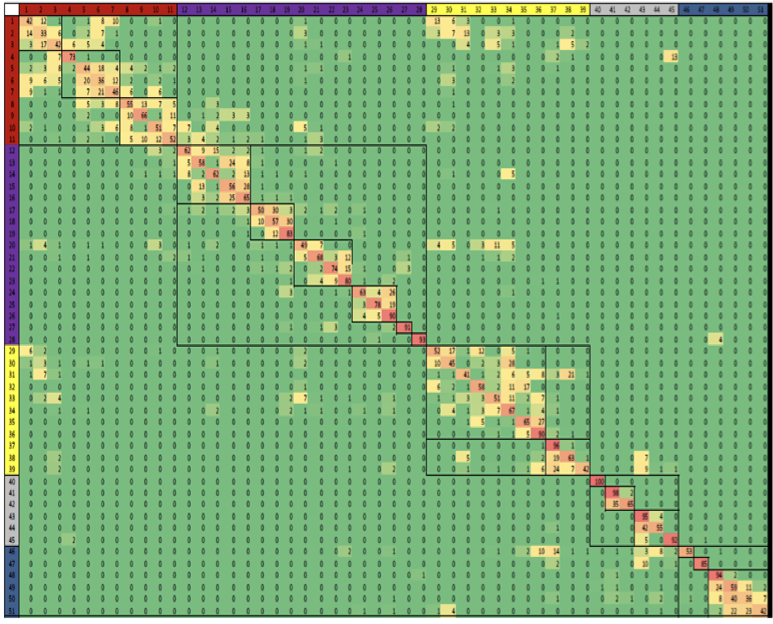
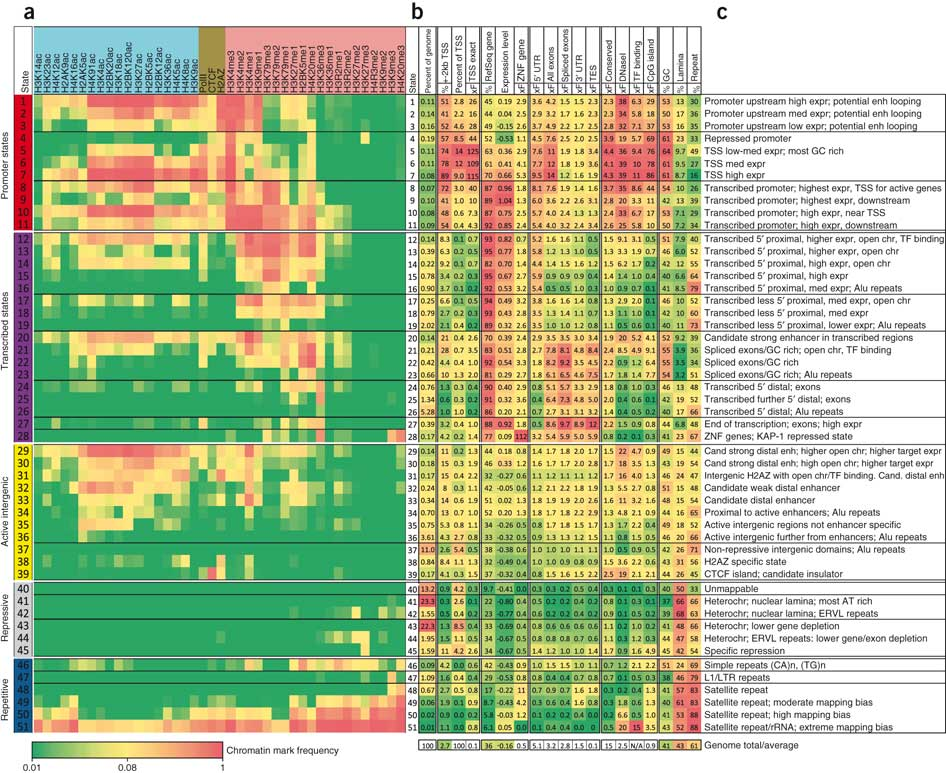
Figura 19.9: Probabilidades de emisión para el modelo final con 51 estados. La celda correspondiente a la marca i y al estado k representa la probabilidad de que la marca i se observe en el estado k.Figura 19.10: Probabilidades de transición para el modelo final con 51 estados. La probabilidad de transición aumenta de verde a rojo. Las relaciones espaciales entre estados de cromatina vecinos y distintos subgrupos de estados se revelan agrupando la matriz de transición. Notablemente, la matriz es escasa, por lo que indica que la mayoría no son posibles.Figura 19.11: Definición del estado de cromatina e interpretación funcional. [7] a. Combinaciones de marcas de cromatina asociadas a cada estado. Cada fila muestra la combinación específica de marcas asociadas a cada estado de cromatina y las frecuencias entre 0 y 1 con las que ocurren en escala de colores. Estos corresponden a los parámetros de probabilidad de emisión del HMM aprendidos a través del genoma durante el entrenamiento del modelo. b. Enriquecimientos genómicos y funcionales de estados de cromatina, incluido el enriquecimiento en veces en la parte dierente del genoma (por ejemplo, regiones transcritas, TSS, RefSeq 5 o 3end del gen, etc.), además plegar el enriquecimiento para elementos conservados evolutivamente, sitios hipersensibles a la ADNasa I, islas CpG, etc. Todos los enriquecimientos se basan en las asignaciones de probabilidad posteriores. c. Breve descripción de la función e interpretación del estado biológico (chr, cromatina; enh, potenciador).