
7.7: Meiosis
- Page ID
- 56667

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)La mitosis produce dos células con el mismo número de cromosomas que la célula parental. La mitosis de una célula diploide (2n) produce dos células hijas diploides. Si dos células diploides continuaran participando en la reproducción sexual, su fusión produciría un cigoto tetraploide (4n). La solución para este problema es la Meiosis.
Meiosis
La meiosis es un proceso de división celular en eucariotas caracterizado por:
- dos divisiones consecutivas: meiosis I y meiosis II
- sin síntesis de ADN (sin fase S) entre las dos divisiones
- el resultado: 4 células con la mitad del número de cromosomas de la célula inicial, por ejemplo, 2n → n
La fusión de dos de estas células produce un cigoto 2n.
Meiosis en Animales
Se utiliza para producir los gametos: esperma y óvulos
Meiosis en Plantas
Se utiliza para producir esporas. Las esporas son el inicio de la generación de gametofitos que, con el tiempo, producirán gametos (por mitosis porque las células de partida ya son haploides).
Meiosis I
La profase de la meiosis I (profase I) es un proceso más elaborado que la profase de la mitosis (y suele tardar mucho más).
Aquí hay una breve descripción del proceso. A continuación se proporciona una vista más detallada.
- Cuando los cromosomas se hacen visibles por primera vez ya están duplicados, habiéndose duplicado cada homólogo durante la fase S anterior.
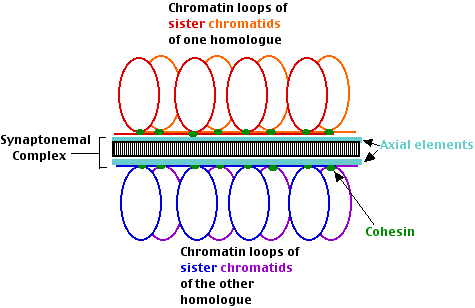
- Resultado: pares de díadas homólogas, cada díada consiste en dos cromátidas hermanas unidas por un complejo proteico llamado cohesina.
- Emparejamiento: Cada par de díadas homólogas se alinean longitudinalmente entre sí.
- Resultado: una tétrada. (Estas estructuras a veces se denominan bivalentes porque en esta etapa no se pueden distinguir las cromátidas hermanas individuales bajo el microscopio).
- Las dos díadas homólogas se mantienen unidas por
- uno o más chiasmata (sing. = quiasma) que se forman entre dos cromátidas no hermanas en puntos donde se han cruzado.
- el complejo sinaptonémico (SC), un complejo ensamblaje de proteínas (incluyendo la cohesina)
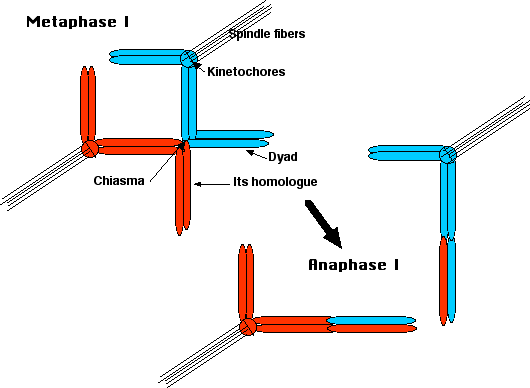
En la metafase I, los microtúbulos de las fibras del huso se adhieren al
- cinetocoros hermanos de un homólogo, tirando de ambas cromátidas hermanas hacia un polo de la célula;
- hermana cinetocoros del otro homólogo tirando a esas hermanas hacia el polo opuesto.
Resultado: se tira de un homólogo por encima de la placa de metafase, el otro debajo. Los chiasmas mantienen a los homólogos unidos entre sí, y el cohesin mantiene unidas a las cromátidas hermanas.
En la anafase I,
- la cohesina entre los brazos cromosómicos se descompone permitiendo
- los chiasmata para que se desplacen.
- Resultado: las díadas homólogas se separan y migran hacia sus respectivos polos.
Meiosis II
El comportamiento cromosómico en la meiosis II es como el de la mitosis.
- En la metafase II, las fibras del huso unen un cinetocoro de la díada a un polo y el otro al polo opuesto.
- En la anafase II, las cromátidas se separan y (cada una ahora es un cromosoma independiente) se mueven a sus respectivos polos.
Recombinación genética
La meiosis no solo preserva el tamaño del genoma de los eucariotas que se reproducen sexualmente, sino que también proporciona tres mecanismos para diversificar los genomas de la descendencia.
Cruce
Los chiasmata representan puntos donde las cromátidas no hermanas anteriores (e invisibles) habían intercambiado secciones. El proceso se llama cruce. Es recíproco; los segmentos intercambiados por cada cromátida no hermana son idénticos (pero pueden llevar diferentes alelos).
Cada cromátida contiene una sola molécula de ADN. Entonces el problema del cruce es realmente un problema de intercambiar porciones de moléculas de ADN adyacentes. Debe hacerse con gran precisión para que ni la cromátida gane ni pierda ningún gen. De hecho, el cruce tiene que ser lo suficientemente preciso para que no se pierda ni se añada ni un solo nucleótido en el punto de cruce si ocurre dentro de un gen. De lo contrario, resultaría un desplazamiento del marco y el gen resultante produciría un producto defectuoso o, más probablemente, ningún producto en absoluto.

En el diagrama anterior, solo se muestra un solo quiasma. Sin embargo, se encuentran comúnmente múltiples chiasmata (en humanos el número promedio de chiasmata por tétrada es poco más de dos). En esta fotomicrografía, una tétrada del saltamontes Chorthippus parallelus muestra 5 chiasmas.
Surtido Aleatorio
En la meiosis I, la orientación de los homólogos paternos y maternos en la placa metafásica es aleatoria. Por lo tanto, aunque cada célula producida por meiosis contiene solo uno de cada homólogo, el número de posibles combinaciones de homólogos maternos y paternos es de 2 n, donde n = el número haploide de cromosomas. En este diagrama, el número haploide es 3, y se producen 8 (2 3) combinaciones diferentes.
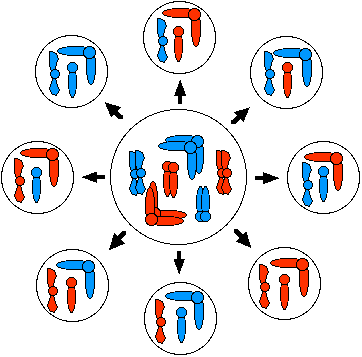
Surtido aleatorio de homólogos en humanos produce 2 23 (8,388,608) combinaciones diferentes de cromosomas.
Además, debido al cruce, ninguno de estos cromosomas es “puro” materno o paterno. La distribución de cromátidas hermanas recombinantes y no recombinantes en las células hijas en la anafase II también es aleatoria.
Entonces creo que es seguro concluir que de todos los miles de millones de espermatozoides producidos por un hombre durante su vida (y los cientos de óvulos que maduran a lo largo de la vida de una mujer), no hay dos que tengan exactamente el mismo contenido genético.
Fertilización
Al reducir el número de cromosomas de 2n a n, se establece la etapa para la unión de dos genomas. Si los padres difieren genéticamente, nuevas combinaciones de genes pueden ocurrir en su descendencia.
Tomando juntos estos tres mecanismos, creo que es seguro concluir que nunca dos seres humanos han compartido un genoma idéntico a menos que hayan tenido un hermano idéntico; es decir, un hermano producido a partir del mismo óvulo fertilizado.
El comportamiento de los cromosomas durante la meiosis (2n → n) y la fertilización (n + n → 2n) proporcionan la base estructural para las reglas de herencia de Mendel.
Profase I: una vista detallada
Los largos y complejos eventos de la profase I pueden dividirse en 5 etapas.
Leptoteno
- Todos los cromosomas se condensan.
- Emparejamiento. Las díadas homólogas (pares de cromátidas hermanas) se encuentran entre sí y se alinean de extremo a extremo con la ayuda de un elemento axial (que contiene cohesina). En la levadura en ciernes (y quizás otros eucariotas) el proceso sigue un periodo de prueba y error. Dos díadas cualesquiera se emparejan en sus centromeros. Si no son homólogos, se separan y vuelven a intentarlo.
- La forma en que las no hermanas reconocen sus regiones compartidas de homología del ADN es incierto. Las roturas bicatenarias (DSB) a menudo ocurren en el ADN de las cromátidas, y estas pueden ser necesarias para que los homólogos se reconozcan entre sí.
Cigoteno
- Sinapsis. El complejo sinaptonémico comienza a formarse.
- Las cadenas de ADN de las cromátidas no hermanas inician el proceso de recombinación. Se desconoce cómo pueden hacerlo a través del complejo sinaptonémico, que tiene más de 100 nm de espesor.
Pachytene
- La sinapsis ya está completa.
- Aparecen nódulos de recombinación (al menos en algunos organismos, incluidos los humanos). Se les llama así por la idea de que representan puntos donde se está produciendo la recombinación de ADN.
- Debe haber al menos uno por cada bivalente para que la meiosis tenga éxito. A menudo hay más, cada uno presumiblemente representa el punto de un crossover.
- Contienen enzimas que se sabe que son necesarias para la recombinación y reparación del ADN.
- Los pasos en la recombinación del ADN continúan hasta el final del paquiteno.
Diploteno
- La recombinación del ADN es completa.
- El complejo sinaptonémico comienza a descomponerse.
- Las cromátidas comienzan a separarse revelando
- chiasmata. Al principio los chiasmas se localizan en los sitios de los nódulos de recombinación, pero posteriormente migran hacia los extremos de las cromátidas.
Diacinesis
En algunos organismos, los cromosomas se descondensan y comienzan a transcribirse por un tiempo. A esto le siguen los cromosomas que se vuelven a condensar en preparación para la metafase I.
En criaturas donde esto no ocurre, los cromosomas se condensan aún más en preparación para la metafase I.
Control de Calidad de Meiosis
No debería sorprender que las cosas puedan salir mal en un proceso tan complicado. Sin embargo, las células que atraviesan meiosis tienen puntos de control que monitorean cada par de homólogos para
- recombinación adecuada de su ADN
- correcta formación del complejo sinaptonémico
Cualquier falla que se detecte detiene el proceso y suele provocar que la célula se autodestruya por apoptosis.
Sin embargo, a pesar de estos puntos de control, los errores ocasionalmente no se corrigen.
Errores en Meiosis
Se estima que del 10 al 25% de todos los óvulos fecundados humanos contienen anomalías cromosómicas, y estas son la causa más común de fracaso del embarazo (35% de los casos).
Estas anomalías cromosómicas
- surgen de errores en la meiosis, generalmente meiosis I;
- ocurren con más frecuencia (90%) durante la formación de óvulos que durante la formación de esperma
- se vuelven más frecuentes a medida que la mujer envejece.
- La aneuploidía —la ganancia o pérdida de cromosomas enteros— es la anomalía cromosómica más común. Es causada por la no disyunción, la falla de los cromosomas para separar correctamente:
- homólogos durante la meiosis I o
- cromátidas hermanas durante la meiosis II
- Los cigotos que carecen de un cromosoma (“monosomía”) no pueden desarrollarse hasta el nacimiento (excepto en las mujeres con un solo cromosoma X).
- Tres del mismo cromosoma (“trisomía”) también son letales a excepción de los cromosomas 13, 18 y 21 (la trisomía 21 es la causa del síndrome de Down).
- Tres o más cromosomas X son viables porque todos menos uno de ellos están inactivados.