
12.2: Determinar las relaciones evolutivas
- Page ID
- 53682

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Los científicos recopilan información que les permite hacer conexiones evolutivas entre organismos. Similar al trabajo de detectives, los científicos deben usar pruebas para descubrir los hechos. En el caso de la filogenia, las investigaciones evolutivas se centran en dos tipos de evidencia: morfológica (forma y función) y genética.
Dos medidas de similitud
Los organismos que comparten características físicas similares y secuencias genéticas tienden a estar más estrechamente relacionados que los que no lo hacen. Las características que se superponen tanto morfológica como genéticamente se denominan estructuras homólogas; las similitudes provienen de caminos evolutivos comunes. Por ejemplo, como se muestra en la Figura\(\PageIndex{1}\), los huesos en las alas de murciélagos y aves, los brazos de los humanos, y la pata delantera de un caballo son estructuras homólogas. Observe que la estructura no es simplemente un solo hueso, sino más bien una agrupación de varios huesos dispuestos de manera similar en cada organismo a pesar de que los elementos de la estructura pueden haber cambiado de forma y tamaño.

Apariciones engañosas
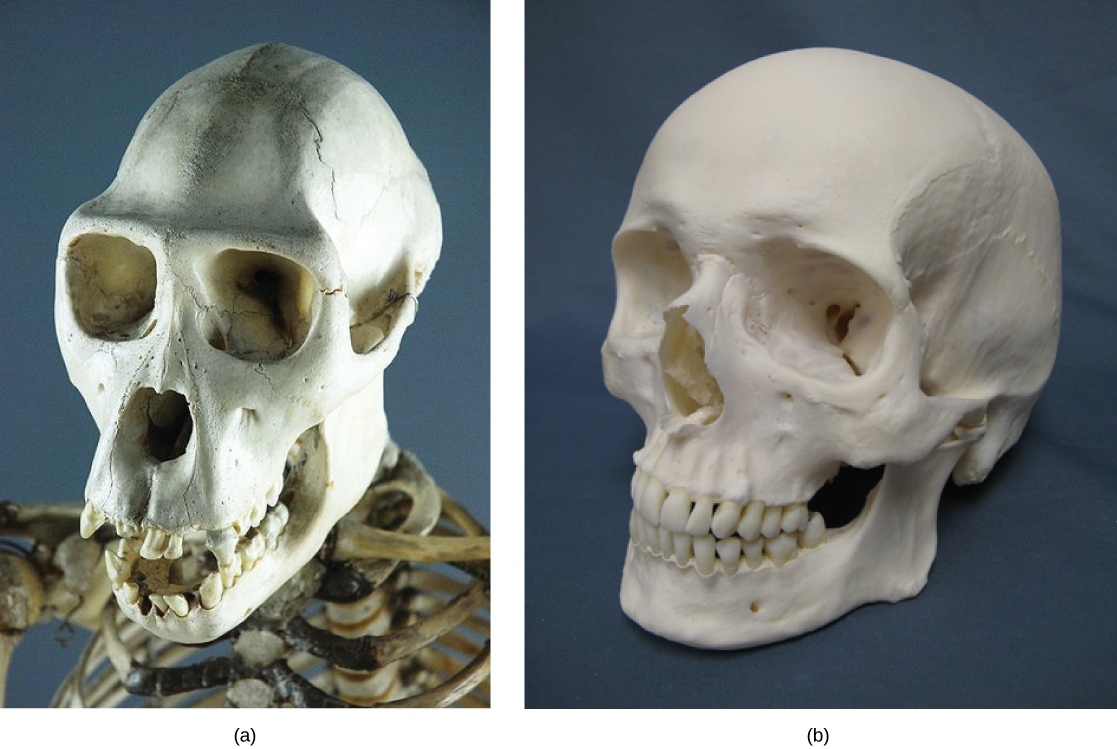
Algunos organismos pueden estar muy estrechamente relacionados, a pesar de que un cambio genético menor provocó una gran diferencia morfológica para que se vieran bastante diferentes. Por ejemplo, los chimpancés y los humanos, cuyos cráneos se muestran en la Figura\(\PageIndex{2}\) son muy similares genéticamente, compartiendo 99 por ciento 1 de sus genes. Sin embargo, los chimpancés y los humanos muestran diferencias anatómicas considerables, incluyendo el grado en que sobresale la mandíbula en el adulto y la longitud relativa de nuestros brazos y piernas.
Sin embargo, los organismos no relacionados pueden estar relacionados de manera lejana pero parecen muy parecidos, generalmente porque las adaptaciones comunes a condiciones ambientales similares evolucionaron en ambos. Un ejemplo son las formas aerodinámicas del cuerpo, las formas de aletas y apéndices, y la forma de las colas en peces y ballenas, que son mamíferos. Estas estructuras tienen similitud superficial porque son adaptaciones al movimiento y maniobra en el mismo ambiente: el agua. Cuando una característica que es similar ocurre por convergencia adaptativa (evolución convergente), y no por una estrecha relación evolutiva, se le llama estructura análoga. En otro ejemplo, los insectos usan alas para volar como murciélagos y aves. Las llamamos ambas alas porque realizan la misma función y tienen una forma superficialmente similar, pero el origen embrionario de las dos alas es completamente diferente. La diferencia en el desarrollo, o embriogénesis, de las alas en cada caso es una señal de que insectos y murciélagos o aves no comparten un ancestro común que tenía un ala. Las estructuras del ala, mostradas en la Figura,\(\PageIndex{3}\) evolucionaron independientemente en los dos linajes.
Rasgos similares pueden ser homólogos o análogos. Los rasgos homólogos comparten un camino evolutivo que condujo al desarrollo de ese rasgo, y los rasgos análogos no. Los científicos deben determinar qué tipo de similitud exhibe una característica para descifrar la filogenia de los organismos estudiados.

Comparaciones Moleculares
Con el avance de la tecnología del ADN, ha florecido el área de la sistemática molecular, que describe el uso de información a nivel molecular incluyendo la secuenciación del ADN. Un nuevo análisis de caracteres moleculares no sólo confirma muchas clasificaciones anteriores, sino que además descubre errores cometidos anteriormente. Los caracteres moleculares pueden incluir diferencias en la secuencia de aminoácidos de una proteína, diferencias en la secuencia de nucleótidos individual de un gen o diferencias en las disposiciones de los genes. Las filogenias basadas en caracteres moleculares suponen que cuanto más similares son las secuencias en dos organismos, más estrechamente relacionadas están. Diferentes genes cambian evolutivamente a diferentes velocidades y esto afecta el nivel en el que son útiles para identificar relaciones. Las secuencias de rápida evolución son útiles para determinar las relaciones entre especies estrechamente relacionadas. Las secuencias que evolucionan más lentamente son útiles para determinar las relaciones entre especies relacionadas distantemente. Para determinar las relaciones entre especies muy diferentes como Eukarya y Archaea, los genes utilizados deben ser genes muy antiguos, que evolucionan lentamente y que están presentes en ambos grupos, como los genes para el ARN ribosómico. Comparar árboles filogenéticos usando diferentes secuencias y encontrarlos similares ayuda a generar confianza en las relaciones inferidas.
En ocasiones, dos segmentos de ADN en organismos distantes relacionados comparten aleatoriamente un alto porcentaje de bases en las mismas localizaciones, lo que hace que estos organismos aparezcan estrechamente relacionados cuando no lo están. Por ejemplo, la mosca de la fruta comparte el 60 por ciento de su ADN con los humanos. 2 En esta situación, se han desarrollado algoritmos estadísticos basados en computadora para ayudar a identificar las relaciones reales y, en última instancia, el uso acoplado de información tanto morfológica como molecular es más efectivo para determinar la filogenia.
EVOLUCIÓN EN ACCIÓN: ¿Por qué importa la filogenia?
Además de mejorar nuestra comprensión de la historia evolutiva de las especies, nuestro propio análisis filogenético incluido tiene numerosas aplicaciones prácticas. Dos de esas aplicaciones incluyen comprender la evolución y transmisión de enfermedades y tomar decisiones sobre los esfuerzos de conservación. Un estudio 3 de 2010 de MRSA (Staphylococcus aureus resistente a meticilina), una bacteria patógena resistente a antibióticos, rastreó el origen y la propagación de la cepa a lo largo de los últimos 40 años. El estudio descubrió el momento y los patrones en los que la cepa resistente se trasladó de su punto de origen en Europa a centros de infección y evolución en América del Sur, Asia, América del Norte y Australasia. El estudio sugiere que las introducciones de la bacteria a nuevas poblaciones ocurrieron muy pocas veces, quizás solo una vez, y luego se propagaron desde ese número limitado de individuos. Esto contrasta con la posibilidad de que muchos individuos hubieran transportado la bacteria de un lugar a otro. Este resultado sugiere que los funcionarios de salud pública deben concentrarse en identificar rápidamente los contactos de individuos infectados con una nueva cepa de bacterias para controlar su propagación.
Una segunda área de utilidad para el análisis filogenético es la conservación. Los biólogos han argumentado que es importante proteger las especies a lo largo de un árbol filogenético en lugar de solo las de una rama del árbol. Hacer esto preservará más de la variación producida por la evolución. Por ejemplo, los esfuerzos de conservación deben centrarse en una sola especie sin especies hermanas en lugar de otra especie que tenga un grupo de especies hermanas cercanas que evolucionaron recientemente. Si la única especie evolutivamente distinta se extingue, se perderá una cantidad desproporcionada de variación del árbol en comparación con una especie en el grupo de especies estrechamente relacionadas. Un estudio publicado en 2007 4 hizo recomendaciones para la conservación de especies de mamíferos en todo el mundo basadas en cuán evolutivamente distintas y en riesgo de extinción son. El estudio encontró que sus recomendaciones diferían de las prioridades basadas simplemente en el nivel de amenaza de extinción para la especie. El estudio recomendó proteger a algunos mamíferos grandes amenazados y valorados como los orangutanes, los pandas gigantes y menores, y los elefantes africanos y asiáticos. Pero también encontraron que algunas especies mucho menos conocidas deberían ser protegidas en función de cuán evolutivas distintas son. Estos incluyen una serie de roedores, murciélagos, musarañas y erizos. Además, hay algunas especies en peligro crítico que no calificaron como muy importantes en la distinción evolutiva, incluyendo especies de ratones venados y jerbos. Si bien muchos criterios afectan las decisiones de conservación, preservar la diversidad filogenética proporciona una manera objetiva de proteger toda la gama de diversidad generada por la evolución.
Construyendo árboles filogenéticos
¿Cómo construyen los científicos árboles filogenéticos? Actualmente, el método más aceptado para construir árboles filogenéticos es un método llamado cladística. Este método clasifica a los organismos en clados, grupos de organismos que están más estrechamente relacionados entre sí y el antepasado del que descienden. Por ejemplo, en la Figura\(\PageIndex{4}\), todos los organismos en la región sombreada evolucionaron a partir de un solo ancestro que tenía huevos amnióticos. En consecuencia, todos estos organismos también tienen huevos amnióticos y forman un solo clado, también llamado grupo monofilético. Los clados deben incluir las especies ancestrales y todos los descendientes de un punto de ramificación.
CONEXIÓN ART
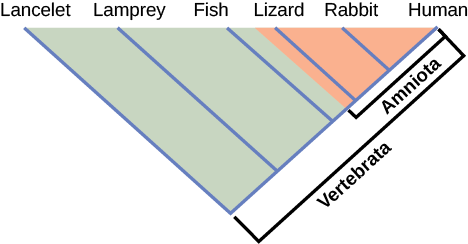
¿Qué animales en esta figura pertenecen a un clado que incluye animales con pelo? ¿Cuál evolucionó primero: el pelo o el óvulo amniótico?
Los clados pueden variar en tamaño dependiendo del punto de ramificación al que se haga referencia. El factor importante es que todos los organismos del clado o grupo monofilético provienen de un solo punto del árbol. Esto se puede recordar porque el monofilético se descompone en “mono”, que significa uno, y “filético”, que significa relación evolutiva.
Características compartidas
La cladística se basa en tres supuestos. La primera es que los seres vivos se relacionan por descendencia de un ancestro común, lo que es una suposición general de evolución. La segunda es que la especiación ocurre por divisiones de una especie en dos, nunca más de dos a la vez, y esencialmente en un punto en el tiempo. Esto es algo polémico, pero es aceptable para la mayoría de los biólogos como simplificación. El tercer supuesto es que los rasgos cambian lo suficiente con el tiempo para ser considerados como en un estado diferente.También se supone que se puede identificar la dirección real del cambio para un estado. En otras palabras, asumimos que un óvulo amniótico es un estado de carácter posterior que los óvulos no amnióticos. A esto se le llama la polaridad del cambio de carácter. Esto lo sabemos por referencia a un grupo fuera del clado: por ejemplo, los insectos tienen huevos no amnióticos; por lo tanto, este es el estado de carácter más antiguo o ancestral. La cladística compara grupos ingroups y outgroups. Un grupo interno (lagarto, conejo y humano en nuestro ejemplo) es el grupo de taxones que se están analizando. Un grupo externo (lanceleta, lamprea y peces en nuestro ejemplo) es una especie o grupo de especies que divergieron antes del linaje que contiene el grupo o grupos de interés. Al comparar los miembros del grupo entre sí y con los miembros del grupo externo, podemos determinar qué características son modificaciones evolutivas determinando los puntos de ramificación de la filogenia del grupo.
Si se encuentra una característica en todos los miembros de un grupo, se trata de un personaje ancestral compartido porque no ha habido cambio en el rasgo durante el descenso de cada uno de los miembros del clado. Si bien estos rasgos parecen interesantes porque unifican el clado, en cladística se consideran no útiles cuando estamos tratando de determinar las relaciones de los miembros del clado porque cada miembro es el mismo. En contraste, considere el óvulo amniótico característico de la Figura\(\PageIndex{4}\). Sólo algunos de los organismos tienen este rasgo, y a los que sí lo hacen, se le llama carácter derivado compartido porque este rasgo cambió en algún momento durante el descenso. Este personaje sí nos habla de las relaciones entre los miembros del clado; nos dice que lagartos, conejos y humanos se agrupan más estrechamente que cualquiera de estos organismos con peces, lampreas y lanceletas.
Un aspecto a veces confuso de los caracteres “ancestrales” y “derivados” es que estos términos son relativos. El mismo rasgo podría ser ancestral o derivado dependiendo del diagrama que se utilice y de los organismos que se comparen. Los científicos encuentran estos términos útiles a la hora de distinguir entre clados durante la construcción de árboles filogenéticos, pero es importante recordar que su significado depende del contexto.
Elegir las relaciones adecuadas
Construir un árbol filogenético, o cladograma, a partir de los datos de los personajes es una tarea monumental que suele dejarse en manos de una computadora. La computadora dibuja un árbol tal que todos los clados comparten la misma lista de caracteres derivados. Pero hay otras decisiones a tomar, por ejemplo, ¿y si la presencia de una especie en un clado es apoyada por todos los personajes derivados compartidos para ese clado excepto uno? Una conclusión es que el rasgo evolucionó en el antepasado, pero luego cambió de nuevo en esa especie. También se debe suponer que un estado de carácter que aparece en dos clados ha evolucionado independientemente en esos clados. Estas inconsistencias son comunes en árboles extraídos de datos de caracteres y complican el proceso de toma de decisiones sobre qué árbol representa más estrechamente las relaciones reales entre los taxones.
Para ayudar en la tremenda tarea de elegir el mejor árbol, los científicos suelen utilizar un concepto llamado parsimonia máxima, lo que significa que los eventos ocurrieron de la manera más simple, más obvia. Esto significa que el “mejor” árbol es el que tiene el menor número de inversiones de carácter, el menor número de cambios de carácter independientes y el menor número de cambios de carácter en todo el árbol. Los programas de computadora buscan a través de todos los árboles posibles para encontrar el pequeño número de árboles con las vías evolutivas más simples. A partir de todos los rasgos homólogos en un grupo de organismos, los científicos pueden determinar el orden de los eventos evolutivos de los que ocurrieron esos rasgos que es el más obvio y sencillo.
CONCEPT EN ACCIÓN

Practica Parsimonia: Ve a este sitio web para conocer cómo se usa la parsimonia máxima para crear árboles filogenéticos (asegúrate de continuar a la segunda página).
Estas herramientas y conceptos son solo algunas de las estrategias que los científicos utilizan para abordar la tarea de revelar la historia evolutiva de la vida en la Tierra. Recientemente, las nuevas tecnologías han descubierto descubrimientos sorprendentes con relaciones inesperadas, como el hecho de que las personas parecen estar más estrechamente relacionadas con los hongos que los hongos con las plantas. ¿Suena increíble? A medida que crece la información sobre las secuencias de ADN, los científicos se acercarán más a mapear la historia evolutiva de toda la vida en la Tierra
Resumen de la Sección
Para construir árboles filogenéticos, los científicos deben recopilar información de carácter que les permita establecer conexiones evolutivas entre organismos. Utilizando datos morfológicos y moleculares, los científicos trabajan para identificar características homólogas y genes. Las similitudes entre organismos pueden provenir de la historia evolutiva compartida (homologías) o de caminos evolutivos separados (analogías). Después de identificar la información homóloga, los científicos utilizan la cladística para organizar estos eventos como un medio para determinar una línea de tiempo evolutiva. Los científicos aplican el concepto de máxima parsimonia, que establece que el orden más probable de eventos es probablemente el camino más corto más simple. Para los eventos evolutivos, este sería el camino con el menor número de divergencias mayores que se correlacionan con la evidencia.
Conexiones de arte
Figura\(\PageIndex{3}\): ¿Qué animales en esta figura pertenecen a un clado que incluye animales con pelo? ¿Cuál evolucionó primero: el pelo o el óvulo amniótico?
- Contestar
-
Conejos y humanos pertenecen al clado que incluye animales con pelo. El huevo amniótico evolucionó antes que el pelo, porque el clado Amniota se ramifica antes que el clado que engloba a los animales con pelo.
Notas al pie
- 1 Gibbons, A. (2012, 13 de junio). Ciencia Ahora. Recuperado de news.sciencemag.org/scienceno... sequenced.html
- 2 Antecedentes sobre el análisis genómico comparativo. (2002, diciembre). Recuperado a partir de http://www.genome.gov/10005835
- 3 Harris, S.R. et al. 2010. Evolución del SARM durante la transmisión hospitalaria y la propagación intercontinental. Ciencia 327:469 —474.
- 4 Isaac NJ, Turvey ST, Collen B, Waterman C, Baillie JE (2007) Mamíferos en el EDGE: Prioridades de conservación basadas en amenazas y filogenia. PLoS ONE 2 (3): e296. doi:10.1371/periodista.pone.0000296
Glosario
- estructura análoga
- un personaje que se encuentra en dos taxones que se parece a causa de la evolución convergente, no por la descendencia de un ancestro común
- clado
- un grupo de taxones con el mismo conjunto de caracteres derivados compartidos, incluyendo una especie ancestral y todos sus descendientes
- cladística
- un método utilizado para organizar rasgos homólogos para describir filogenias usando el descendiente común como criterio principal utilizado para clasificar organismos
- parsimonia máxima
- aplicando la forma más simple y obvia con el menor número de pasos
- sistemática molecular
- los métodos de uso de evidencia molecular para identificar relaciones filogenéticas
- grupo monofilético
- (también, clado) organismos que comparten un solo ancestro
- carácter ancestral compartido
- un personaje en una rama filogenética que es compartida por un clado particular
- carácter derivado compartido
- un personaje en un árbol filogenético que es compartido solo por cierto clado de organismos