
27.1: La crisis de la biodiversidad
- Page ID
- 58252
Tradicionalmente, los ecologistas han medido la biodiversidad, término general para la variedad presente en la biosfera, tomando en cuenta tanto el número de especies como su comunalidad. La biodiversidad se puede estimar en varios niveles de organización de los seres vivos. Estos índices de estimación, que provienen de la teoría de la información, son más útiles como primer paso para cuantificar la biodiversidad entre y dentro de los ecosistemas; son menos útiles cuando la principal preocupación entre los biólogos de conservación es simplemente la pérdida de biodiversidad. Sin embargo, los biólogos reconocen que las medidas de biodiversidad, en términos de diversidad de especies, pueden ayudar a enfocar los esfuerzos para preservar los elementos biológica o tecnológicamente importantes de la biodiversidad.
Los cíclidos del lago Victoria proporcionan un ejemplo a través del cual podemos comenzar a comprender la biodiversidad. Los biólogos que estudiaron cíclidos en la década de 1980 descubrieron cientos de especies de cíclidos que representan una variedad de especializaciones para tipos de hábitat particulares y estrategias de alimentación específicas: comer plancton flotando en el agua, raspando y luego comiendo algas de las rocas, comiendo larvas de insectos desde el fondo, y comer los huevos de otras especies de cíclidos. Los cíclidos del lago Victoria son producto de una radiación adaptativa. Una radiación adaptativa es una rápida ramificación (menos de tres millones de años en el caso de los cíclidos del lago Victoria) a través de la especiación de un árbol filogenético en muchas especies estrechamente relacionadas; típicamente, la especie “irradia” a diferentes hábitats y nichos. Los pinzones de Galápagos son un ejemplo de una modesta radiación adaptativa con 15 especies. Los cíclidos del lago Victoria son un ejemplo de una espectacular radiación adaptativa que incluye alrededor de 500 especies.
En el momento en que los biólogos realizaban este descubrimiento, algunas especies comenzaron a desaparecer rápidamente. Un culpable de estos descensos fue una especie de peces grandes que fue introducida en el lago Victoria por la pesca para alimentar a las personas que viven alrededor del lago. La perca del Nilo se introdujo en 1963, pero permaneció baja hasta la década de 1980 cuando sus poblaciones comenzaron a crecer. La población de percas del Nilo creció consumiendo cíclidos, llevando especies tras especies al punto de extinción (la desaparición de una especie). De hecho, hubo varios factores que jugaron un papel en la extinción de quizás 200 especies de cíclidos en el lago Victoria: la perca del Nilo, la disminución de la calidad del agua del lago debido a la agricultura y la limpieza de tierras a orillas del lago Victoria, y el aumento de la presión pesquera. Los científicos ni siquiera habían catalogado todas las especies presentes, tantas se perdieron que nunca fueron nombradas. La diversidad es ahora una sombra de lo que alguna vez fue.
Los cíclidos del lago Victoria son un boceto en miniatura de la rápida pérdida de especies contemporáneas que ocurre en toda la Tierra y es causada por la actividad humana. La extinción es un proceso natural de macroevolución que ocurre a razón de que aproximadamente una de cada 1 millón de especies se extingue por año. El registro fósil revela que ha habido cinco períodos de extinción masiva en la historia con tasas mucho más altas de pérdida de especies, y la tasa de pérdida de especies hoy en día es comparable a esos períodos de extinción masiva. Sin embargo, existe una gran diferencia entre las extinciones masivas anteriores y la extinción actual que estamos viviendo: la actividad humana. Específicamente, tres actividades humanas tienen un impacto importante: la destrucción del hábitat, la introducción de especies exóticas y la sobrecosecha. Predicciones de pérdida de especies dentro del próximo siglo, una pequeña cantidad de tiempo en escalas de tiempo geológicas, que van del 10 por ciento al 50 por ciento. Las extinciones en esta escala sólo han ocurrido otras cinco veces en la historia del planeta, y han sido causadas por eventos cataclísmicos que cambiaron el curso de la historia de la vida en cada instancia. Muchos científicos creen que la Tierra está entrando ahora en una sexta extinción masiva.
Tipos de Biodiversidad
Los científicos generalmente aceptan que el término biodiversidad describe el número y los tipos de especies en un lugar o en el planeta. Las especies pueden ser difíciles de definir, pero la mayoría de los biólogos aún se sienten cómodos con el concepto y son capaces de identificar y contar especies eucariotas en la mayoría de los contextos. Los biólogos también han identificado medidas alternas de biodiversidad, algunas de las cuales son importantes para planificar cómo preservar la biodiversidad.
La diversidad genética es uno de esos conceptos alternos. La diversidad o variación genética es la materia prima para la adaptación en una especie. El potencial futuro de adaptación de una especie depende de la diversidad genética que se mantenga en los genomas de los individuos en las poblaciones que componen la especie. Lo mismo es cierto para categorías taxonómicas superiores. Un género con tipos muy diferentes de especies tendrá más diversidad genética que un género con especies que se parecen y tienen ecologías similares. Si hubiera una elección entre uno de estos géneros de especies que se conservaba, el que tiene mayor potencial para su posterior evolución es el más genéticamente diverso. Sería ideal no tener que tomar tales decisiones, pero cada vez más esta puede ser la norma.
Muchos genes codifican proteínas, que a su vez llevan a cabo los procesos metabólicos que mantienen vivos y reproduciéndose a los organismos. La diversidad genética se puede medir como diversidad química en que diferentes especies producen una variedad de químicos en sus células, tanto las proteínas como los productos y subproductos del metabolismo. Esta diversidad química tiene beneficios potenciales para los humanos como fuente de productos farmacéuticos, por lo que proporciona una forma de medir la diversidad que es importante para la salud y el bienestar humanos.
Los humanos han generado diversidad en animales domésticos, plantas y hongos. Esta diversidad también está sufriendo pérdidas debido a la migración, las fuerzas del mercado y el aumento del globalismo en la agricultura, especialmente en regiones densamente pobladas como China, India y Japón. La población humana depende directamente de esta diversidad como fuente estable de alimento, y su declive es preocupante para biólogos y científicos agrícolas.
También es útil para definir la diversidad de ecosistemas, es decir, el número de ecosistemas diferentes en el planeta o en un área geográfica determinada. Los ecosistemas enteros pueden desaparecer aunque algunas de las especies sobrevivan adaptándose a otros ecosistemas. La pérdida de un ecosistema significa la pérdida de interacciones entre especies, la pérdida de características únicas de la coadaptación y la pérdida de productividad biológica que un ecosistema es capaz de crear. Un ejemplo de un ecosistema en gran parte extinto en América del Norte es el ecosistema de las praderas. Las praderas alguna vez abarcaron América Central del Norte desde el bosque boreal en el norte de Canadá hasta México. Ahora están casi desaparecidas, reemplazadas por campos de cultivo, pastos y expansión suburbana. Muchas de las especies sobreviven, pero el ecosistema enormemente productivo que se encargó de crear los suelos agrícolas más productivos ya no está. Como consecuencia, los suelos están desapareciendo o deben mantenerse a mayor costo.
Diversidad Actual de Especies
A pesar de un esfuerzo considerable, el conocimiento de las especies que habitan el planeta es limitado. Una estimación reciente sugiere que las especies eucariotas para las que la ciencia tiene nombres, alrededor de 1.5 millones de especies, representan menos del 20 por ciento del número total de especies eucariotas presentes en el planeta (8.7 millones de especies, por una estimación). Las estimaciones del número de especies procariotas son en gran parte conjeturas, pero los biólogos coinciden en que la ciencia sólo ha comenzado a catalogar su diversidad. Incluso con lo que se sabe, no existe un repositorio central de nombres o muestras de las especies descritas; por lo tanto, no hay forma de estar seguro de que los 1.5 millones de descripciones sean un número exacto. Es una mejor conjetura basada en las opiniones de expertos en diferentes grupos taxonómicos. Dado que la Tierra está perdiendo especies a un ritmo acelerado, la ciencia está muy en el lugar que estaba con los cíclidos del lago Victoria: saber poco sobre lo que se está perdiendo. \(\PageIndex{1}\)En la tabla se presentan estimaciones recientes de biodiversidad en diferentes grupos.
| Tabla\(\PageIndex{1}\): Estimaciones de los números de especies descritas y predichas por grupo taxonómico | ||||||
|---|---|---|---|---|---|---|
| Mora et al. 2011 | Chapman | Groombridge & Jenkins 2002 | ||||
| \ (\ PageIndex {1}\): Estimaciones de los números de especies descritas y predichas por grupo taxonómico"> | \ (\ PageIndex {1}\): Estimaciones de los números de especies descritas y predichas por grupo taxonómicoMora et al. 2011">Descrito | Predijo | \ (\ PageIndex {1}\): Estimaciones de los números de especies descritas y predichas por grupo taxonómicoChapman 2009">Descrito | Predijo | \ (\ PageIndex {1}\): Estimaciones de los números de especies descritas y predichas por grupo taxonómicoGroomBridge & Jenkins 2002">Descrito | Predijo |
| \ (\ PageIndex {1}\): Estimaciones del número de especies descritas y predichas por grupo taxonómico">Animalia | \ (\ PageIndex {1}\): Estimaciones de los números de especies descritas y predichas por grupo taxonómicoMora et al. 2011">1,124,516 | 9,920,000 | \ (\ PageIndex {1}\): Estimaciones de los números de especies descritas y predichas por grupo taxonómicoChapman 2009">1,424,153 | 6,836,330 | \ (\ PageIndex {1}\): Estimaciones de los números de especies descritas y predichas por grupo taxonómicoGroombridge & Jenkins 2002">1,225,500 | 10,820,000 |
| \ (\ PageIndex {1}\): Estimaciones de los números de especies descritas y predichas por grupo taxonómico">Chromista | \ (\ PageIndex {1}\): Estimaciones de los números de especies descritas y predichas por grupo taxonómicoMora et al. 2011">17.892 | 34,900 | \ (\ PageIndex {1}\): Estimaciones de los números de especies descritas y predichas por grupo taxonómicoChapman 2009">25.044 | 200,500 | \ (\ PageIndex {1}\): Estimaciones de los números de especies descritas y predichas por grupo taxonómicoGroomBridge & Jenkins 2002">— | — |
| \ (\ PageIndex {1}\): Estimaciones del número de especies descritas y predichas por grupo taxonómico">Hongos | \ (\ PageIndex {1}\): Estimaciones de los números de especies descritas y predichas por grupo taxonómicoMora et al. 2011">44.368 | 616,320 | \ (\ PageIndex {1}\): Estimaciones de los números de especies descritas y predichas por grupo taxonómicoChapman 2009">98,998 | 1,500,000 | \ (\ PageIndex {1}\): Estimaciones de los números de especies descritas y predichas por grupo taxonómicoGroombridge & Jenkins 2002">72,000 | 1,500,000 |
| \ (\ PageIndex {1}\): Estimaciones de los números de especies descritas y predichas por grupo taxonómico">Plantae | \ (\ PageIndex {1}\): Estimaciones de los números de especies descritas y predichas por grupo taxonómicoMora et al. 2011">224.244 | 314,600 | \ (\ PageIndex {1}\): Estimaciones de los números de especies descritas y predichas por grupo taxonómicoChapman 2009">310,129 | 390,800 | \ (\ PageIndex {1}\): Estimaciones de los números de especies descritas y predichas por grupo taxonómicoGroombridge & Jenkins 2002">270,000 | 320,000 |
| \ (\ PageIndex {1}\): Estimaciones del número de especies descritas y predichas por grupo taxonómico">Protozoos | \ (\ PageIndex {1}\): Estimaciones de los números de especies descritas y predichas por grupo taxonómicoMora et al. 2011">16.236 | 72,800 | \ (\ PageIndex {1}\): Estimaciones de los números de especies descritas y predichas por grupo taxonómicoChapman 2009">28,871 | 1,000,000 | \ (\ PageIndex {1}\): Estimaciones de los números de especies descritas y predichas por grupo taxonómicoGroombridge & Jenkins 2002">80.000 | 600,000 |
| \ (\ PageIndex {1}\): Estimaciones del número de especies descritas y predichas por grupo taxonómico">Procariotas | \ (\ PageIndex {1}\): Estimaciones de los números de especies descritas y predichas por grupo taxonómicoMora et al. 2011">— | — | \ (\ PageIndex {1}\): Estimaciones de los números de especies descritas y predichas por grupo taxonómicoChapman 2009">10.307 | 1,000,000 | \ (\ PageIndex {1}\): Estimaciones de los números de especies descritas y predichas por grupo taxonómicoGroombridge & Jenkins 2002">10,175 | — |
| \ (\ PageIndex {1}\): Estimaciones de los números de especies descritas y predichas por grupo taxonómico">Total | \ (\ PageIndex {1}\): Estimaciones de los números de especies descritas y predichas por grupo taxonómicoMora et al. 2011">1,438.769 | 10,960,000 | \ (\ PageIndex {1}\): Estimaciones de los números de especies descritas y predichas por grupo taxonómicoChapman 2009">1,897,502 | 10,897,630 | \ (\ PageIndex {1}\): Estimaciones de los números de especies descritas y predichas por grupo taxonómicoGroombridge & Jenkins 2002">1.657,675 | 13,240,000 |
Existen diversas iniciativas para catalogar especies descritas de manera accesible, e internet está facilitando ese esfuerzo. No obstante, se ha señalado que al ritmo actual de descripción de especies, que según el Informe del Estado de las Especies Observadas es de 17,000 a 20,000 nuevas especies por año, tardará cerca de 500 años en terminar de describir la vida en este planeta (IISE, 2011). Con el tiempo, la tarea se vuelve cada vez más imposible y cada vez más fácil a medida que la extinción elimina especies del planeta.
Nombrar y contar especies puede parecer una búsqueda sin importancia dadas las otras necesidades de la humanidad, pero no es simplemente una contabilidad. Describir especies es un proceso complejo mediante el cual los biólogos determinan las características únicas de un organismo y si ese organismo pertenece o no a alguna otra especie descrita. Permite a los biólogos encontrar y reconocer la especie después del descubrimiento inicial y les permite dar seguimiento a preguntas sobre su biología. Además, las características únicas de cada especie la hacen potencialmente valiosa para los humanos u otras especies de las que dependen los humanos. Comprender estas características es el valor de encontrar y nombrar especies.
Patrones de Biodiversidad
La biodiversidad no se distribuye uniformemente en la Tierra. El lago Victoria solo contenía casi 500 especies de cíclidos, ignorando las otras familias de peces presentes en el lago. Todas estas especies se encontraron solo en el lago Victoria; por lo tanto, las 500 especies de cíclidos fueron endémicas. Las especies endémicas se encuentran en un solo lugar. Los endémicos con distribuciones altamente restringidas son particularmente vulnerables a la extinción. Los niveles taxonómicos más altos, como géneros y familias, también pueden ser endémicos. El lago Huron contiene alrededor de 79 especies de peces, todas las cuales se encuentran en muchos otros lagos de América del Norte. ¿Qué explica la diferencia en la diversidad de peces en estos dos lagos? El lago Victoria es un lago tropical, mientras que el lago Huron es un lago templado. El lago Huron en su forma actual tiene apenas unos 7.000 años, mientras que el Lago Victoria en su forma actual tiene unos 15 mil años. Los biogeógrafos han sugerido que estos dos factores, latitud y edad, son dos de varias hipótesis para explicar los patrones de biodiversidad en el planeta.
La biogeografía es el estudio de la distribución de las especies del mundo, tanto en el pasado como en el presente. El trabajo de los biogeógrafos es fundamental para comprender nuestro entorno físico, cómo el ambiente afecta a las especies y cómo los cambios ambientales impactan la distribución de una especie; también ha sido crítico para desarrollar la teoría evolutiva. Los biogeógrafos necesitan entender tanto la biología como la ecología. También necesitan estar bien versados en estudios evolutivos, ciencia del suelo y climatología.
Hay tres campos principales de estudio bajo el rubro de biogeografía: biogeografía ecológica, biogeografía histórica (llamada paleobiogeografía) y biogeografía de conservación. La biogeografía ecológica estudia los factores actuales que afectan la distribución de plantas y animales. La biogeografía histórica, como su nombre lo indica, estudia la distribución pasada de las especies. La biogeografía de conservación, por otro lado, se enfoca en la protección y restauración de especies con base en información ecológica histórica y actual conocida. Cada uno de estos campos considera tanto la zoogeografía como la fitogeografía, la distribución pasada y presente de animales y plantas.
Uno de los patrones más antiguos observados en ecología es que la biodiversidad de especies en casi todos los grupos taxonómicos aumenta a medida que disminuye la latitud. Es decir, la biodiversidad aumenta más cerca del ecuador (Figura\(\PageIndex{1}\)).
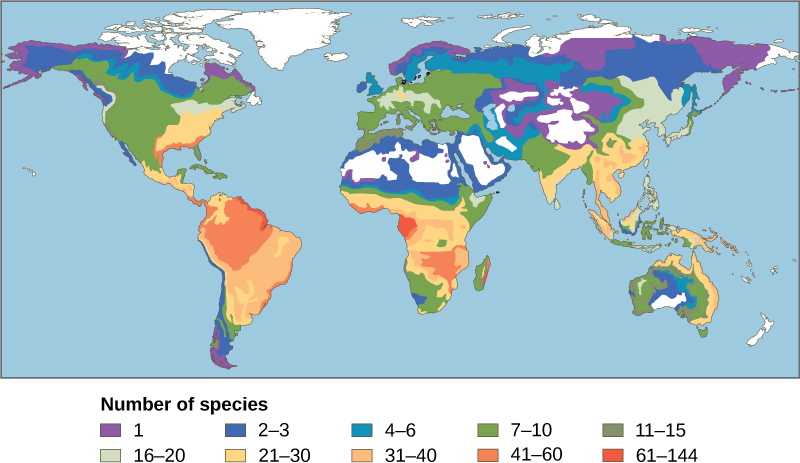
Aún no está claro por qué la biodiversidad aumenta más cerca del ecuador, pero las hipótesis incluyen la mayor edad de los ecosistemas en los trópicos versus regiones templadas que estuvieron en gran parte desprovistas de vida o drásticamente empobrecidas durante la última glaciación. La idea es que una mayor edad proporcione más tiempo para la especiación. Otra posible explicación es el aumento de la energía que reciben los trópicos del sol frente a la disminución de la energía que reciben las regiones templadas y polares. No está del todo claro cómo un mayor aporte de energía podría traducirse en más especies. La complejidad de los ecosistemas tropicales puede promover la especiación al aumentar la heterogeneidad, o número de nichos ecológicos, en los trópicos en relación con latitudes más altas. La mayor heterogeneidad brinda más oportunidades de coevolución, especialización y quizás mayores presiones de selección que conducen a la diferenciación poblacional. Sin embargo, esta hipótesis adolece de cierta circularidad: los ecosistemas con más especies fomentan la especiación, pero ¿cómo consiguieron más especies para empezar? Los trópicos han sido percibidos como más estables que las regiones templadas, las cuales presentan un clima pronunciado y una estacionalidad diurna pronunciada. Los trópicos tienen sus propias formas de estacionalidad, como la lluvia, pero generalmente se supone que son ambientes más estables y esta estabilidad podría promover la especiación.
Independientemente de los mecanismos, sin duda es cierto que todos los niveles de biodiversidad son mayores en los trópicos. Adicionalmente, la tasa de endemismo es la más alta, y hay más puntos críticos de biodiversidad. Sin embargo, esta riqueza de diversidad también significa que nuestro conocimiento de las especies es más bajo, y existe un alto potencial de pérdida de biodiversidad.
Conservación de la Biodiversidad
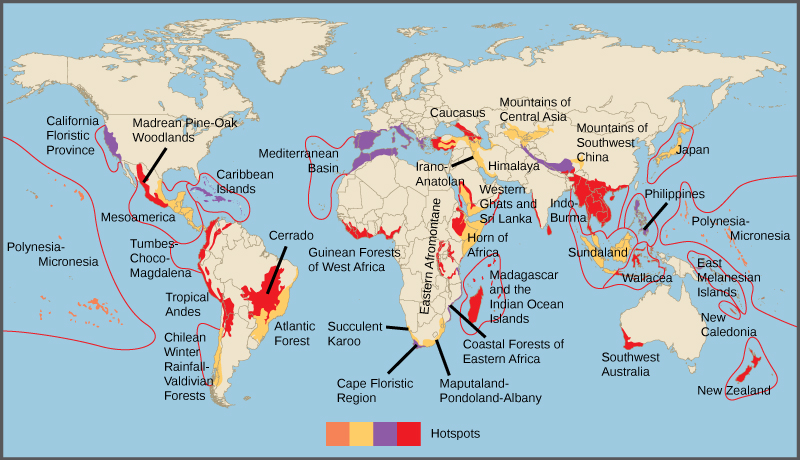
En 1988, el ambientalista británico Norman Myers desarrolló un concepto de conservación para identificar áreas ricas en especies y con riesgo significativo de pérdida de especies: los puntos críticos de biodiversidad. Los puntos calientes de biodiversidad son áreas geográficas que contienen un alto número de especies endémicas. El propósito del concepto fue identificar ubicaciones importantes en el planeta para los esfuerzos de conservación, una especie de triaje de conservación. Al proteger los puntos calientes, los gobiernos son capaces de proteger a un mayor número de especies. Los criterios originales para un hotspot incluyeron la presencia de 1500 o más especies de plantas endémicas y 70 por ciento del área perturbada por la actividad humana. Actualmente hay 34 puntos calientes de biodiversidad (Figura\(\PageIndex{2}\)) que contienen un gran número de especies endémicas, que incluyen la mitad de las plantas endémicas de la Tierra.
Cambio de biodiversidad a través del tiempo geológico
El número de especies en el planeta, o en cualquier área geográfica, es el resultado de un equilibrio de dos procesos evolutivos que están en curso: la especiación y la extinción. Ambos son procesos naturales de “nacimiento” y “muerte” de macroevolución. Cuando las tasas de especiación comiencen a superar las tasas de extinción, el número de especies aumentará; asimismo, el número de especies disminuirá cuando las tasas de extinción comiencen a superar a las tasas de especiación. A lo largo de la historia de la Tierra, estos dos procesos han fluctuado, a veces conduciendo a cambios dramáticos en el número de especies en la Tierra como se refleja en el registro fósil (Figura\(\PageIndex{3}\)).
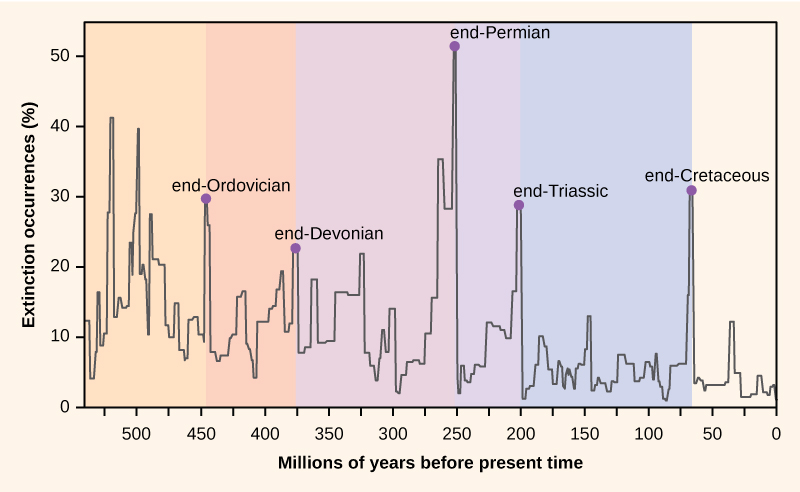
Los paleontólogos han identificado cinco estratos en el registro fósil que parecen mostrar pérdidas repentinas y dramáticas (más de la mitad de todas las especies existentes que desaparecen del registro fósil) en la biodiversidad. A estas se les llama extinciones masivas. Hay muchos eventos de extinción menores, pero aún dramáticos, pero las cinco extinciones masivas han atraído la mayor cantidad de investigación. Se puede argumentar que las cinco extinciones masivas son sólo los cinco eventos más extremos en una serie continua de grandes eventos de extinción a lo largo del Fanerozoico (desde hace 542 millones de años). En la mayoría de los casos, las causas hipotéticas siguen siendo polémicas.
Las cinco extinciones masivas
El registro fósil de las extinciones masivas fue la base para definir períodos de la historia geológica, por lo que suelen ocurrir en el punto de transición entre períodos geológicos. La transición en los fósiles de un período a otro refleja la dramática pérdida de especies y el origen gradual de nuevas especies. Estas transiciones se pueden ver en los estratos rocosos. La tabla\(\PageIndex{2}\) proporciona datos sobre las cinco extinciones masivas.
| Extinciones Masivas | ||
|---|---|---|
| Periodo Geológico | Nombre de extinción masiva | Tiempo (hace millones de años) |
| Ordovícico-Silúrico | O—S de extremo ordovícico | 450—440 |
| Devónico tardío | Final-devónico | 375—360 |
| Pérmico-Triásico | Pérmico final | 251 |
| Triásico-Jurásico | Final-triásico | 205 |
| Cretácico-Paleógeno | Cretácico Final K—Pg (K—T) | 65.5 |
El evento de extinción Ordovícico-Silúrico es la primera extinción masiva registrada y la segunda más grande. Durante este periodo, alrededor del 85 por ciento de las especies marinas (pocas especies vivían fuera de los océanos) se extinguieron. La hipótesis principal para su causa es un período de glaciación y luego calentamiento. El evento de extinción en realidad consiste en dos eventos de extinción separados por alrededor de 1 millón de años. El primer evento fue causado por el enfriamiento, y el segundo evento se debió al posterior calentamiento. Los cambios climáticos afectaron las temperaturas y el nivel del mar. Algunos investigadores han sugerido que una explosión de rayos gamma, causada por una supernova cercana, es una posible causa de la extinción Ordovicia-Silúrico. El estallido de rayos gamma habría despojado la capa de ozono de la Tierra provocando una intensa radiación ultravioleta del sol y podría explicar los cambios climáticos observados en ese momento. La hipótesis es especulativa, pero las influencias extraterrestres en la historia de la Tierra son una línea activa de investigación. La recuperación de la biodiversidad después de la extinción masiva tomó de 5 a 20 millones de años, dependiendo de la ubicación.
La extinción tardía del Devónico puede haber ocurrido durante un período de tiempo relativamente largo. Parece haber afectado a especies marinas y no a las plantas o animales que habitan hábitats terrestres. Las causas de esta extinción son poco conocidas.
La extinción final del Pérmico fue la más grande en la historia de la vida. En efecto, se podría argumentar que la Tierra casi se volvió desprovista de vida durante este suceso de extinción. El planeta se veía muy diferente antes y después de este evento. Se estima que el 96 por ciento de todas las especies marinas y el 70 por ciento de todas las especies terrestres se perdieron. Fue en este momento, por ejemplo, que los trilobites, grupo que sobrevivió a la extinción Ordovícico-Silúrico, se extinguieron. Las causas de esta extinción masiva no están claras, pero el principal sospechoso es la actividad volcánica extendida y generalizada que condujo a un evento desbocado de calentamiento global. Los océanos se volvieron en gran parte anóxicos, sofocando la vida marina. La diversidad de tetrápodos terrestres tardó 30 millones de años en recuperarse tras la extinción final del Pérmico. La extinción Pérmica alteró drásticamente la composición de la biodiversidad de la Tierra y el curso de la evolución.
Las causas del evento de extinción Triásico-Jurásico no están claras y se han argumentado hipótesis sobre el cambio climático, el impacto de asteroides y las erupciones volcánicas. El evento de extinción ocurrió justo antes de la desintegración del supercontinente Pangea, aunque recientes estudios sugieren que la extinción puede haber ocurrido de manera más gradual a lo largo del Triásico.
Las causas del evento de extinción del Cretácico Final son las que mejor se entienden. Fue durante este suceso de extinción hace unos 65 millones de años que los dinosaurios, el grupo vertebrado dominante durante millones de años, desaparecieron del planeta (con la excepción de un clado terópodo que dio origen a aves). Cada animal terrestre que pesaba más de 25 kg se extinguió. Ahora se entiende que la causa de esta extinción es el resultado de un impacto cataclísmico de un gran meteorito, o asteroide, frente a la costa de lo que hoy es la Península de Yucatán. Esta hipótesis, propuesta por primera vez en 1980, fue una explicación radical basada en un fuerte pico en los niveles de iridio (que llueve desde el espacio en meteoros a un ritmo bastante constante pero que por lo demás está ausente en la superficie de la Tierra) en el estrato rocoso que marca el límite entre los períodos Cretácico y Paleógeno (Figura\(\PageIndex{4}\)). Este límite marcó la desaparición de los dinosaurios tanto en fósiles como en muchos otros taxones. Los investigadores que descubrieron el pico de iridio lo interpretaron como una rápida afluencia de iridio del espacio a la atmósfera (en forma de un gran asteroide) en lugar de una ralentización en la deposición de sedimentos durante ese período. Fue una explicación radical, pero el reporte de un cráter de impacto apropiadamente envejecido y dimensionado en 1991 hizo que la hipótesis fuera más creíble. Ahora una abundancia de evidencia geológica respalda la teoría. Los tiempos de recuperación de la biodiversidad después de la extinción del Cretácico Final son más cortos, en tiempo geológico, que para la extinción final del Pérmico, del orden de 10 millones de años.

La extinción del Pleistoceno
La Extinción del Pleistoceno es una de las extinciones menores y una reciente. Es bien sabido que los norteamericanos, y hasta cierto punto eurasiáticos, megafauna, o animales grandes, desaparecieron hacia el final del último período de glaciación. La extinción parece haber ocurrido en un periodo de tiempo relativamente restringido de hace 10 mil a 12 mil años. En Norteamérica, las pérdidas fueron bastante dramáticas e incluyeron a los mamuts lanudos (fechados por última vez hace unos 4.000 años en una población aislada), mastodontes, castores gigantes, perezosos gigantes de tierra, dientes de sable gatos, y el camello norteamericano, sólo por nombrar algunos. La posibilidad de que la rápida extinción de estos grandes animales fuera causada por la caza excesiva se sugirió por primera vez en la década de 1900. La investigación sobre esta hipótesis continúa hoy en día. Parece probable que la caza excesiva haya causado muchas extinciones de historia preescritas en muchas regiones del mundo.
En general, el momento de las extinciones del Pleistoceno se correlacionó con la llegada de los humanos y no con los eventos del cambio climático, que es la principal hipótesis competidora para estas extinciones. Las extinciones comenzaron en Australia hace unos 40 mil a 50 mil años, justo después de la llegada de los humanos a la zona: desaparecieron un león marsupial, un wombat gigante de una tonelada y varias especies de canguros gigantes. En América del Norte, la extinción de casi todos los grandes mamíferos ocurrió hace 10 mil a 12 mil años. Todo lo que queda son los mamíferos más pequeños como osos, alces, alces y pumas. Finalmente, en muchas islas oceánicas remotas, la extinción de muchas especies ocurrió coincidente con las llegadas de humanos. No todas las islas tenían animales grandes, pero cuando había animales grandes, se perdieron. Madagascar fue colonizado hace unos 2 mil años y los grandes mamíferos que allí vivían se extinguieron. Eurasia y África no muestran este patrón, pero tampoco experimentaron una reciente llegada de humanos. Los humanos llegaron a Eurasia hace cientos de miles de años después del origen de la especie en África. Este tema sigue siendo un área de investigación activa y de hipótesis. Parece claro que aunque el clima jugara un papel, en la mayoría de los casos la caza humana precipitó las extinciones.
Extinciones en Tiempo Presente
La sexta extinción masiva (también llamada extinción del Holoceno) parece haber comenzado antes de lo que se creía anteriormente y tiene que ver principalmente con las actividades del Homo sapiens. Desde el inicio del período Holoceno, existen numerosas extinciones recientes de especies individuales que se registran en escritos humanos. La mayoría de estos son coincidentes con la expansión de las colonias europeas desde el siglo XVI.
Uno de los ejemplos anteriores y popularmente conocidos es el ave dodo. El pájaro dodo vivía en los bosques de Mauricio, una isla en el Océano Índico. El pájaro dodo se extinguió alrededor de 1662. Fue cazada por su carne por marineros y era presa fácil porque el dodo, que no evolucionaba con los humanos, se acercaría a la gente sin miedo. Los cerdos, ratas y perros introducidos traídos a la isla por barcos europeos también mataron a dodo jóvenes y huevos.
La vaca marina de Steller se extinguió en 1768; estaba relacionada con el manatí y probablemente alguna vez vivió a lo largo de la costa noroeste de América del Norte. La vaca marina de Steller fue descubierta por primera vez por los europeos en 1741 y fue cazada en busca de carne y aceite. La última vaca marina fue asesinada en 1768. Eso equivale a 27 años entre el primer contacto de la vaca marina con los europeos y la extinción de la especie.
En 1914, la última paloma pasajera viva murió en un zoológico de Cincinnati, Ohio. Esta especie alguna vez había oscurecido los cielos de América del Norte durante sus migraciones, pero fue cazada y sufrió pérdida de hábitat a través de la tala de bosques para tierras de cultivo. En 1918, el último periquito viviente de Carolina murió en cautiverio. Esta especie alguna vez fue común en el este de Estados Unidos, pero sufrió pérdida de hábitat. La especie también fue cazada porque comió frutos de huerto cuando sus alimentos nativos fueron destruidos para dar paso a tierras de cultivo. El león marino japonés, que habitaba una amplia zona alrededor de Japón y la costa de Corea, se extinguió en la década de 1950 debido a los pescadores. La foca monje caribeña se distribuyó por todo el Mar Caribe pero fue impulsada a la extinción mediante la caza en 1952.
Estas son sólo algunas de las extinciones registradas en los últimos 500 años. La Unión Internacional para la Conservación de la Naturaleza (UICN) mantiene una lista de especies extintas y en peligro de extinción llamada Lista Roja. El listado no está completo, pero describe 380 especies extintas de vertebrados después del 1500 d.C., 86 de las cuales fueron expulsadas por la sobrecaza o sobrepesca.
Estimaciones de tasas de extinción en tiempo actual
Las estimaciones de las tasas de extinción se ven obstaculizadas por el hecho de que la mayoría de las extinciones probablemente ocurren sin observación. Es probable que los humanos noten la extinción de un ave o mamífero, especialmente si ha sido cazada o utilizada de alguna otra manera. Pero hay muchos organismos que son de menor interés para los humanos (no necesariamente de menor valor) y muchos que no están descritos.
La tasa de extinción de fondo se estima en alrededor de una por millón de especies por año (E/MSY). Por ejemplo, suponiendo que existan alrededor de diez millones de especies, la expectativa es que diez especies se extingan cada año (cada año representa diez millones de especies por año).
Una estimación de la tasa de extinción contemporánea utiliza las extinciones en el registro escrito desde el año 1500. Solo para las aves, este método arroja una estimación de 26 E/MSY. Sin embargo, este valor puede ser subestimado por tres razones. En primer lugar, muchas especies no habrían sido descritas hasta mucho más tarde en el periodo de tiempo, por lo que su pérdida habría pasado desapercibida. En segundo lugar, el número de especies recientemente extintas está aumentando debido a que actualmente se describen especies extintas a partir de restos óseos. Y tercero, algunas especies probablemente ya están extintas a pesar de que los conservacionistas son reacios a nombrarlas como tales. Al tomar en cuenta estos factores, la tasa estimada de extinción se aproxima a los 100 E/MSY. La tasa prevista para finales de siglo es 1500 E/MSY.
Un segundo enfoque para estimar las tasas de extinción en tiempo presente es correlacionar la pérdida de especies con la pérdida de hábitat midiendo la pérdida de área forestal y entendiendo las relaciones especie-área. La relación especie-área es la tasa a la que se observan nuevas especies cuando se incrementa el área encuestada. Los estudios han demostrado que el número de especies presentes aumenta a medida que aumenta el tamaño de la isla. También se ha demostrado que este fenómeno se mantiene cierto en otros hábitats. Dando la vuelta a esta relación, si se reduce el área de hábitat, también disminuirá el número de especies que allí habitan. Las estimaciones de las tasas de extinción basadas en la pérdida de hábitat y las relaciones especie-área han sugerido que con alrededor del 90 por ciento de pérdida de hábitat se espera que 50 por ciento de las especies se extinga. Las estimaciones del área de especies han llevado a cálculos de la tasa de extinción de especies de alrededor de 1000 E/MSY y superiores. En general, las observaciones reales no muestran esta cantidad de pérdida y se han hecho sugerencias de que hay un retraso en la extinción. Trabajos recientes también han puesto en tela de juicio la aplicabilidad de la relación especie-área al estimar la pérdida de especies. En este trabajo se argumenta que la relación especie-área conduce a una sobreestimación de las tasas de extinción. Una mejor relación para usar puede ser la relación endémico-área. El uso de este método reduciría las estimaciones a alrededor de 500 E/MSY en el próximo siglo. Tenga en cuenta que este valor sigue siendo 500 veces la tasa de fondo.
Resumen
La biodiversidad existe en múltiples niveles de organización y se mide de diferentes maneras dependiendo de los objetivos de quienes toman las medidas. Estas mediciones incluyen números de especies, diversidad genética, diversidad química y diversidad de ecosistemas. El número de especies descritas se estima en 1.5 millones con alrededor de 17,000 especies nuevas que se describen cada año. Las estimaciones para el número total de especies en la Tierra varían pero son del orden de 10 millones. La biodiversidad se correlaciona negativamente con la latitud para la mayoría de los taxones, lo que significa que la biodiversidad es mayor en los trópicos. El mecanismo para este patrón no se conoce con certeza, pero se han avanzado varias hipótesis plausibles.
Cinco extinciones masivas con pérdidas de más del 50 por ciento de las especies existentes son observables en el registro fósil. Los tiempos de recuperación de la biodiversidad después de las extinciones masivas varían, pero han sido de hasta 30 millones de años. Las extinciones recientes se registran en la historia escrita y son la base de un método de estimación de las tasas de extinción contemporáneas. El otro método utiliza medidas de pérdida de hábitat y relaciones especie-área. Las estimaciones de las tasas de extinción contemporáneas varían, pero algunas tasas son tan altas como 500 veces la tasa de fondo, según se determina a partir del registro fósil, y se prevé que aumenten.
Referencias
Camilo, M. et al. 2011. “¿Cuántas especies hay en la Tierra y en el Océano?” PLoS Biology, doi:10.1371/journal.pbio.1001127.
Chapman, A. 2009. Números de Especies Vivas en Australia y el Mundo, 2a ed. (Canberra, AU: Estudio Australiano de Recursos Biológicos). http://www.environment.gov.au/biodiv...d-complete.pdf.
Groombridge, B. y Jenkins, M. 2002. Atlas Mundial de Biodiversidad: Recursos Vivos de la Tierra en el Siglo XXI. Berkeley: Prensa de la Universidad de California.
Instituto Internacional de Exploración de Especies (IISE), 2011. Estado de las especies observadas (SOS). Tempe, AZ: IISE, 2011. Consultado el 20 de mayo de 2012. http://species.asu.edu/SOS.
OpenStax, Biología. OpenStax CNX. 26 de junio de 2020. https://cnx.org/contents/GFy_h8cu@10.137:noBcfThl@7/Understanding-Evolution.