
2.5: Gen y Operón
- Page ID
- 57326

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Genes
La secuencia completa de ácido nucleico que es necesaria para la síntesis de un polipéptido funcional o molécula de ARN. Así, un gen contiene información de secuencia adicional más allá de la que codifica para los aminoácidos en una proteína o los nucleótidos en una molécula de ARN. El gen también contiene el ADN necesario para obtener una transcripción particular hecha.
Las regiones de control de la transcripción pueden estar alejadas de la región codificante (en el orden de Kb o 10 de Kb de distancia).
La mayoría de los genes procariotas carecen de intrones (secuencia de ADN intercalada). En los procariotas, los genes que codifican proteínas con relaciones en una vía metabólica forman Operones, que producen ARNm policistrónicos.
Definiciones: Operón y Promotor
- Un operón se encuentra en el ADN bacteriano, un grupo de genes contiguos transcritos a partir de un promotor que da lugar a un ARNm policistrónico.
- Un promotor es una secuencia de ADN a la que se une la ARN polimerasa antes del inicio de la transcripción, generalmente se encuentra justo aguas arriba del sitio de inicio de la transcripción de un gen.
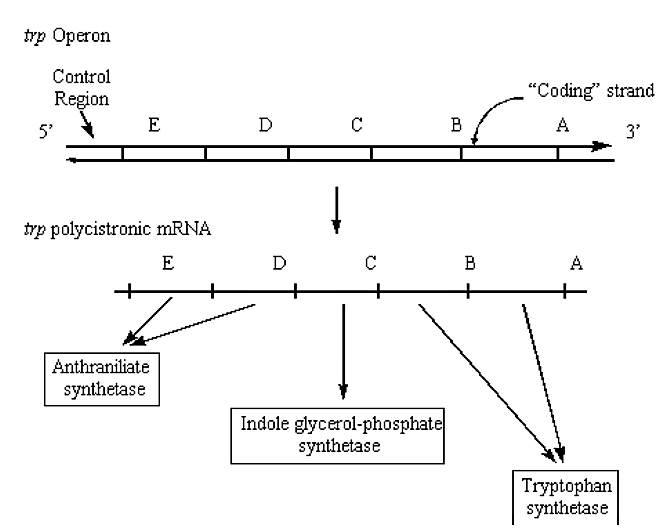
ej., Operon Trp - implicado en la biosíntesis del aminoácido triptófano:
.png)
Figura 2.5.1: Vía química del operón trp
.png)
Figura 2.5.2: Operón Trp en ADN/ARN
Una consecuencia de la disposición de los genes bacterianos en operones es que el nivel de ARNm para cada uno de los genes en el operón es exactamente el mismo.
Los ribosomas se transcriben desde el inicio de cada gen, no sólo desde el primer gen.
Otra consecuencia de la disposición de los genes bacterianos en operones es que una mutación aguas arriba (es decir, posiblemente inhibiendo la transcripción) puede evitar que los genes “aguas abajo” sean transcritos y expresados. La mayoría de las unidades de transcripción eucariotas producen ARNm monocistrónicos, (es decir, codifican solo una proteína). Existe una diferencia fundamental en los procesos de traducción de procariotas y eucariotas:
- En procariotas, los ribosomas pueden unirse a secuencias de reconocimiento específicas en cualquier lugar dentro del ARNm (llamados sitios de unión a ribosomas, o sitios “Shine-Dalgarno”).
- En eucariotas, los ribosomas se unen a través de la interacción con la región 5' específicamente modificada (el llamado sitio 5' cap) de las moléculas de ARNm.
- Por lo tanto, la mayoría de los ARNm eucariotas son monocistrónicos.
- Las mutaciones en unidades de transcripción eucariotas simples afectan solo a una proteína.
Unidades de transcripción eucariotas complejas
El transcrito de ARN primario codificado por unidades de transcripción complejas se puede empalmar de más de una manera. Debido a las diferentes posibilidades de procesamiento, los exones (regiones codificantes) en una sola unidad de transcripción compleja se pueden unir de formas alternativas, para producir diferentes ARNm y diferentes proteínas.
.png)
Figura 2.5.3: Unidades de transcripción complejas
Regulación transcripcional
La supervivencia exitosa requiere adaptabilidad y economía:
- La capacidad de cambiar de metabolizar un sustrato a otro a medida que cambian los recursos ambientales
- Sería un desperdicio energético producir enzimas para una vía metabólica que no es necesaria.
Inducción versus represión de síntesis enzimática
En E. coli ciertas enzimas se producen sólo cuando las células se cultivan sobre ciertos sustratos. Este efecto se llama inducción enzimática. Por ejemplo, cuando las células se cultivan en ausencia de un tipo de azúcar conocido como b galacósido (por ejemplo lactosa) las células contienen muy pocas moléculas (~5 por célula) de la enzima b-galactosidasa (que escinde la lactosa en glucosa y galactosa).
- No hay necesidad de esta enzima en ausencia de lactosa.
- Si se agrega lactosa a E. coli, en muy poco tiempo hay aproximadamente 5000 moléculas de b -galactosidasa por célula (aproximadamente ~1,000 veces de inducción).
- Si se elimina la lactosa del medio se detiene la síntesis de b -galactosidasa.
Una situación similar pero opuesta ocurre con respecto a la síntesis de triptófano (las enzimas biosintéticas están contenidas en el operón trp). En este caso la producción de las enzimas para la biosíntesis de triptófano se detiene rápidamente si el triptófano está presente, en un proceso llamado represión. La represión es un mecanismo regulador transcripcional para los productos génicos comúnmente requeridos. La inducción es un mecanismo regulador transcripcional para productos génicos que pueden ser requeridos en situaciones inusuales o poco frecuentes.