
9.5: Genética no mendeliana
- Page ID
- 56302

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Codominancia y múltiples alelos
Se dice que la co-dominancia ocurre cuando hay una expresión de dos alelos dominantes. El caso prototípico para esto es el grupo sanguíneo ABO humano.

Existen tres alelos en el sistema ABO: A, B y O. Esto da como resultado cuatro tipos de sangre: A, B, O y el AB mezclado.

Dominio Incompleto
Durante la época de Mendel, las personas creían en un concepto de herencia mixta mediante el cual la descendencia demostraba fenotipos intermedios entre los de la generación parental. Esto fue refutado por los experimentos con guisantes de Mendel que ilustraron una Ley de Dominancia. A pesar de esto, la herencia no mendeliana se puede observar en sexo-vinculación y co-dominancia donde no se observan claramente las proporciones esperadas de fenotipos. El dominio incompleto se asemeja superficialmente a la idea de mezclar la herencia, pero aún se puede explicar usando las leyes de Mendel con modificación. En este caso, los alelos no ejercen dominancia total y las crías se asemejan a una mezcla de los dos fenotipos.
El dominio incompleto en las flores de boca de dragón aparece superficialmente como herencia de mezcla. Crédito: Jeremy Seto (CC-BY-NC-SA)
El caso más obvio de un sistema de dos alelos que exhibe un dominio incompleto es en la flor de boca de dragón. Los alelos que dan origen a la coloración floral (Rojo o Blanco) ambos expresan y el genotipo heterocigótico produce flores rosadas. Hay diferentes formas de denotar esto. En este caso, los superíndices de R o W se refieren a los alelos rojos o blancos, respectivamente. Dado que no existe una dominancia clara, el uso de una letra compartida para denotar el rasgo común con los superíndices (o subíndices) permite una denotación más clara de las traducciones finales de genotipo a fenotipo.
Problema: Dominio Incompleto
Si las flores rosadas surgieran de la herencia mixta, entonces los cruces posteriores de flores rosadas con cualquiera de las cepas parentales continuarían diluyendo el fenotipo. Utilizando un Cuadrado Punnet, se realiza un cruce de prueba entre una planta heterocigota y una parental para predecir los fenotipos de la descendencia.
Epistasis y genes modificadores:

La interacción de múltiples enzimas en una vía bioquímica alterará el fenotipo. Algunos genes modificarán las acciones de otro gen.
Crédito: Jeremy Seto (CC0)
Los genes no existen aislados y los productos génicos a menudo interactúan de alguna manera. Epistasis se refiere al evento en el que un gen en un locus depende de la expresión de un gen en otro locus genómico. Dicho de otra manera, un locus genético actúa como modificador de otro. Esto se puede visualizar fácilmente en el caso de la coloración del labrador retriever donde existen tres esquemas de coloración de la capa primaria: laboratorio negro, laboratorio de chocolate y laboratorio amarillo.

Laboratorio de chocolate (arriba), Laboratorio negro (medio), Laboratorio amarillo (abajo) coloraciones de la capa surgen de la interacción de 2 loci génicos, cada uno con 2 alelos. Crédito: Erikeltic [CC-BY-SA 3.0]
Dos genes están involucrados en la coloración de los labradores. El primero es un gen para una proteína llamada TYRP1, que se localiza en los melanosomas (orgánulos de almacenamiento de pigmentos). Se han identificado tres alelos mutantes de este gen que reducen la función de la proteína y producen una coloración más clara. Estos tres alelos pueden señalarse como “b” mientras que el alelo funcional se llama “B”. Un individuo heterocigótico (Bb) o homocigótico dominante estará recubierto de negro mientras que un individuo homocigótico recesivo (bb) será marrón.

Laboratorio negro (BB o Bb) y Laboratorio de chocolate (bb). Crédito: dmealiffe [CC BY-SA 2.0]
El segundo gen está ligado al gen para el Receptor de Melanocortina 1 (MC1R) e influye si el pigmento de eumelanina se expresa en el fur. Este gen tiene los alelos denotados “E” o “e”. Un labrador amarillo tendrá un genotipo de Bbee o bbee.

Laboratorio negro (EE o Ee) y laboratorio amarillo (ee) [CC0]
La interacción entre estos genes puede describirse mediante el siguiente diagrama:
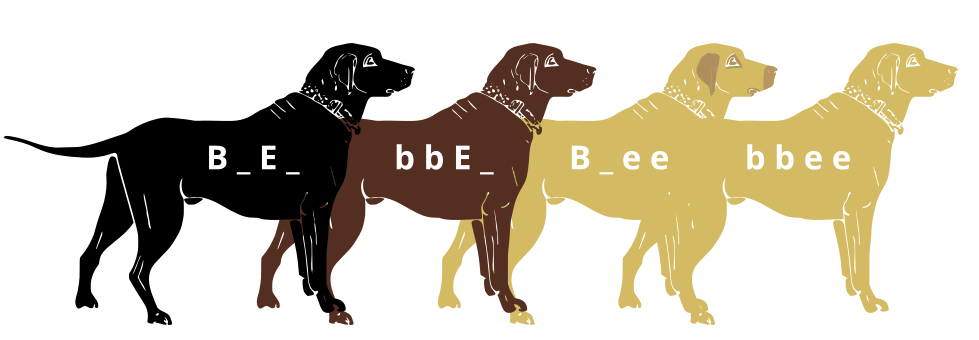
Laboratorio negro (B_E_, Laboratorio de chocolate (BBE_), Laboratorio amarillo con piel oscura donde se expuso (B _ee) y laboratorio amarillo con piel clara donde se expuso. Crédito: Jeremy Seto (CC-BY-SA 3.0)