
12.2: Linaje Materno
- Page ID
- 56381

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Herencia mitocondrial y Materna
Además de los 23 cromosomas heredados de la madre y los 23 cromosomas heredados del padre, los humanos tienen un genoma adicional que sólo se hereda de la madre. Este genoma proviene del orgánulo endosimbiótico, la mitocondria.
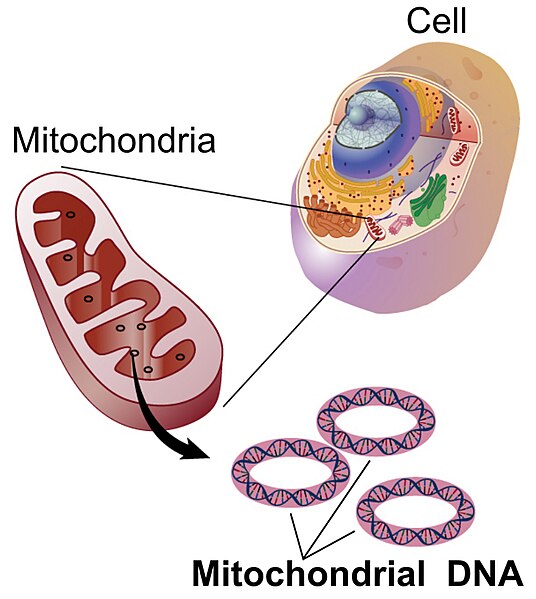
Se cree que las mitocondrias surgieron en la línea eucariota cuando bacterias capaces de desintoxicar los efectos mortales del oxígeno atmosférico fueron absorbidas por un eucariota que no procedió a consumirlo. Con el transcurso del tiempo, estas bacterias que antes vivían libres se volvieron dependientes del entorno celular eucariota mientras proporcionaban el beneficio a la célula hospedadora de la respiración aeróbica. Las características de este evento endosimbiótico incluyen la membrana procariota interna rodeada por la membrana eucariota externa, la presencia de ribosomas procariotas y lo más significativo, y el cromosoma procariótico circular. Las mitocondrias aún se replican independientemente de la célula huésped pero no pueden sobrevivir fuera de este entorno celular. Las mitocondrias animales tienen los genomas más simples de todos los genomas mitocondriales, que van desde 11-28kb. El genoma mitocondrial humano consta de 37 genes que están casi todos dedicados al procesamiento del ATP a través de la fosforilación oxidativa.
Genoma mitocondrial humano
El genoma mitocondrial humano (archivo genbank) consta de 16.569 nucleótidos (16.6kb). Si bien la mayor parte de este genoma de 16.6kb consiste en genes que codifican proteínas, aproximadamente el ADN no codificante de 1.2kb participa en señales que controlan la expresión de estos genes y procesos de replicación. Es el área de ADN donde se desplaza la doble cadena y que tiene el nombre de bucle D (bucle de desplazamiento). Las mutaciones en esta área generalmente tienen muy poco efecto sobre el funcionamiento de las mitocondrias. Debido a esta presión de selección reducida en esta área, esta región de control también se conoce como la región hipervariable. Esta región hipervariable en realidad tiene 10 veces más SNP que el genoma nuclear. Debido a esta abundancia de mutaciones, es posible rastrear la línea materna de un individuo. ¿Por qué solo maternal? El ovocito humano contiene muchas mitocondrias mientras que los espermatozoides solo contienen mitocondrias que potencian el movimiento flagelar. Tras la fecundación, se pierden el flagelo y las mitocondrias asociadas, dejando el cigoto con solo mitocondrias maternas.
El cúmulo de SNP que se encuentra en la región mitocondrial de control están vinculados y siempre se heredan juntos. Debido a la falta de contribución paterna, esta vinculación es referida como un haplotipo, o “medio tipo”. Rastreando estos haplotipos polimórficos, en la década de 1980 se desarrolló un árbol genealógico de humanos que concluyó que los humanos surgieron de una metafórica “Eva Mitocondrial” hace 200 mil años. Como metáfora de la Eva bíblica, esto alude a un origen pero a diferencia del acontecimiento bíblico, esto no quiere decir que fue una sola mujer la que dio origen a toda la humanidad moderna. Por el contrario, la metáfora se limita a indicar que una serie de hembras; hermanas y primos, de esta línea dieron origen a los humanos modernos.

Mapa migratorio de haplogrupos mitocondriales. Los números representan hace 1000 años.
El uso de mitocondrias para este análisis proporciona una gran flexibilidad, especialmente de fuentes antiguas. A diferencia del genoma nuclear que solo tiene 2 copias de ADN por célula, las mitocondrias son abundantes en número y proporcionan muchas copias de genoma por célula. Las fuentes antiguas de ADN en los fósiles suelen tener degradación del ADN. El genoma mitocondrial es igual de probable que se degrada con el tiempo. Sin embargo, el alto número de copias permite rellenar fácilmente los huecos. Los SNP no alteran el tamaño global de la región hipervariable, por lo que la amplificación por PCR no puede resolver estas diferencias con base en la migración del gel de agarosa. Sin embargo, se pueden enviar mplicones (copias amplificadas) para la secuenciación por lo que cada nucleótido puede ser llamado sucesivamente y revelar los SNP específicos.