7.4: Coníferas
- Page ID
- 56368

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Coníferas
Las coníferas son el linaje más rico en especies de gimnospermas. A partir del registro fósil, pensamos que había más de 20 mil especies de coníferas. Sin embargo, su diversidad disminuyó con los dinosaurios. Actualmente, hay alrededor de 600 especies existentes. Estas increíbles plantas representan algunos de los organismos más antiguos, altos y masivos del planeta. Aunque actualmente son bajas en diversidad, estas increíbles plantas constituyen el 30% de los bosques de la Tierra. Las coníferas comparten las siguientes características:
- Monoica. Las plantas producen estrobili tanto machos como hembras en la misma planta.
- Polinizado por el viento con polen “alado” (sacos de aire)
- Hojas xerofíticas con una relación de área superficial a volumen baja. Principalmente de hoja perenne, pero algunas especies son caducifolias (ex. Secoya del amanecer y alerce).
Nota: La Pinaceae es actualmente la familia más grande de coníferas, por lo que muchos de nuestros ejemplos para este grupo de gimnospermas serán del género tipo Pinus (pinos).
Semillas y Polen
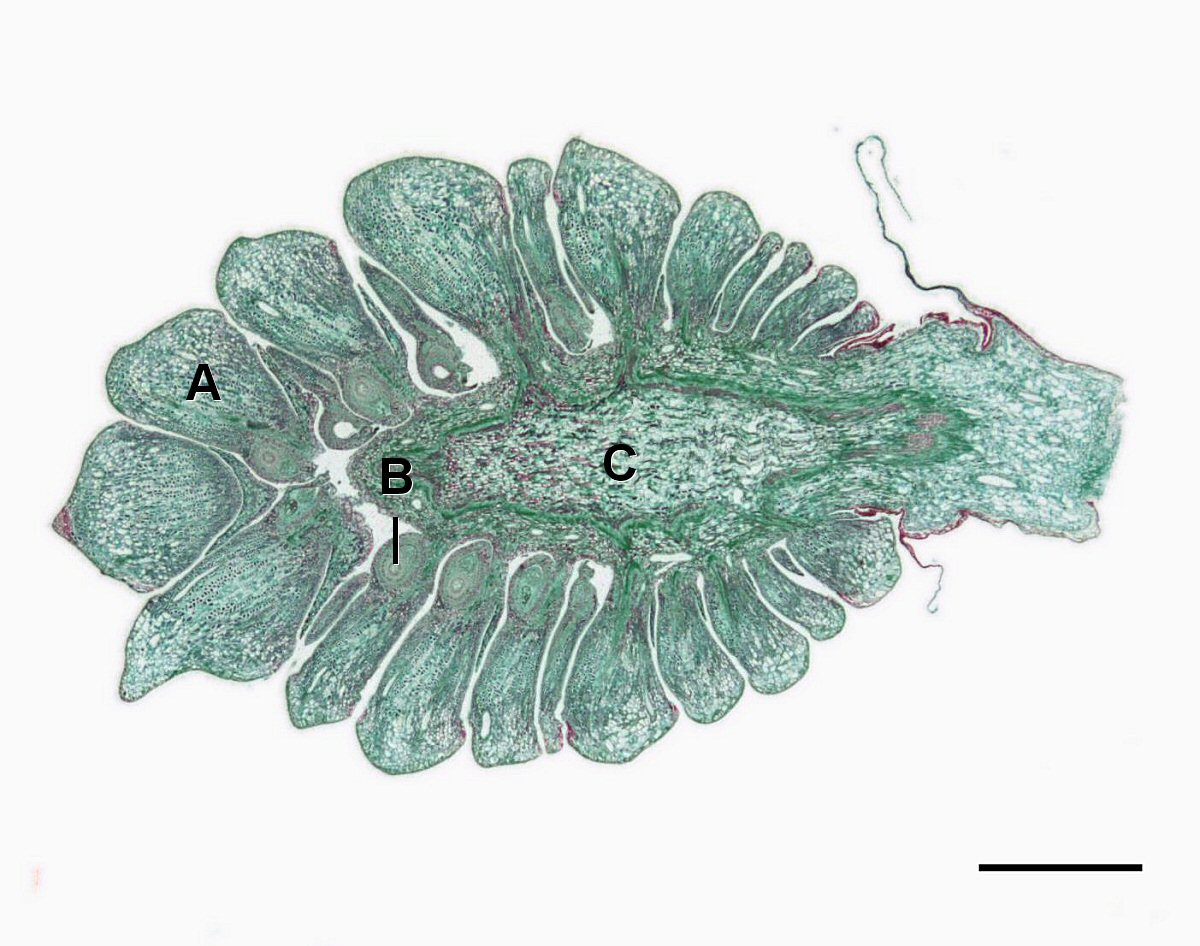
Conos de Semilla


El megastrobilus, o cono semilla, contiene megasporocitos diploides que se producen dentro de un megasporangium.
Cada megasporocito (también llamado célula madre de megasporo) sufre meiosis. Sólo una de las cuatro células producidas sobrevivirá para convertirse en un megagametófito y las otras tres morirán.
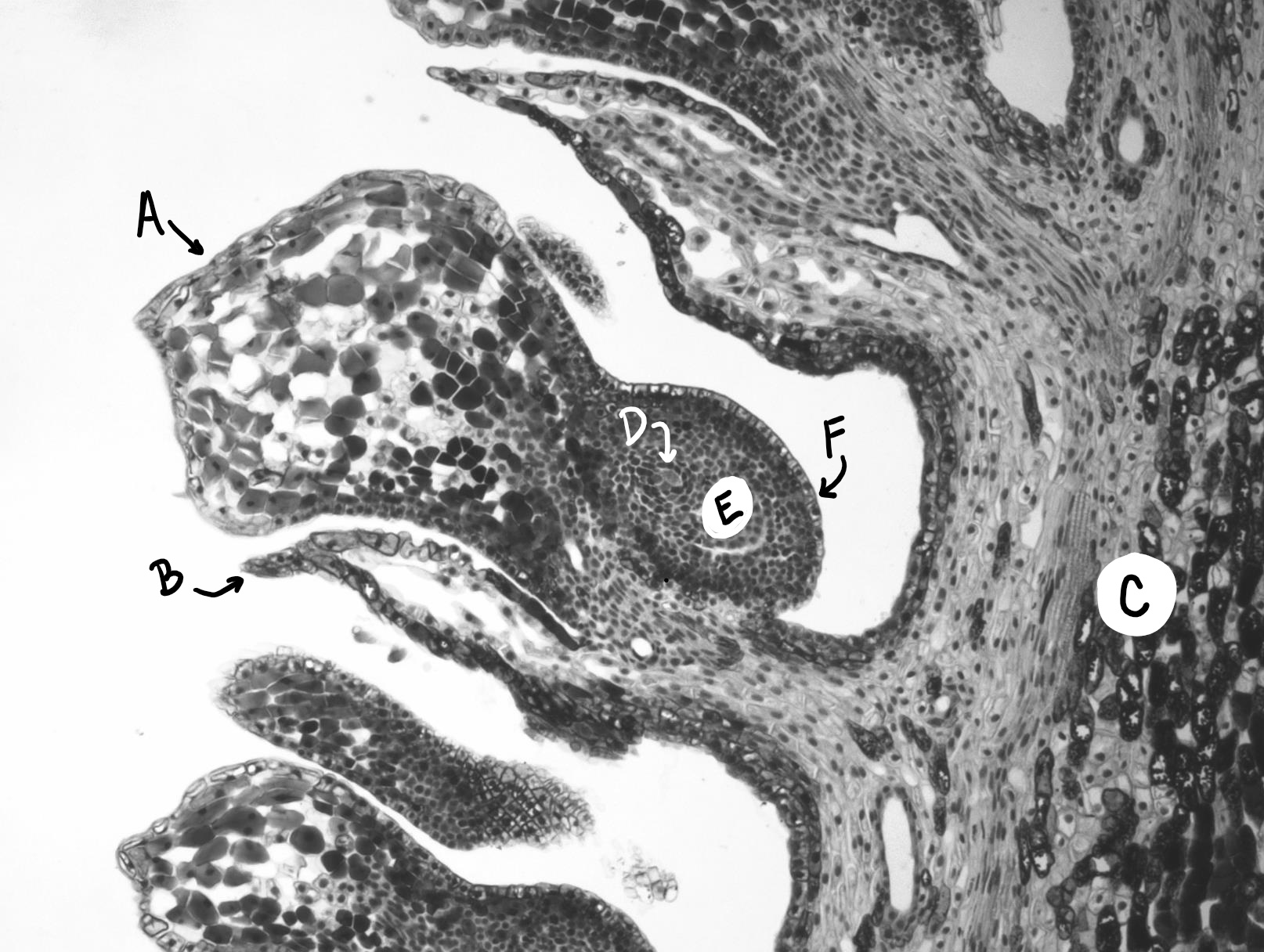
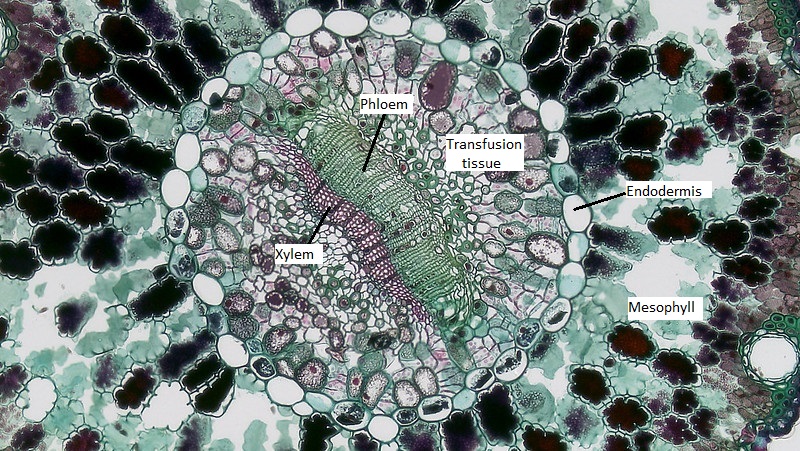
El megagametofito es parte del óvulo y contiene archegonia, cada una con un óvulo en su interior. El megagametofito se retiene dentro del megasporangium, el cual se convierte en el nucello. Rodeando al nucello se encuentra el tegumento, que inicialmente es continuo con la escala ovífera y tiene una pequeña abertura llamada micropilo.
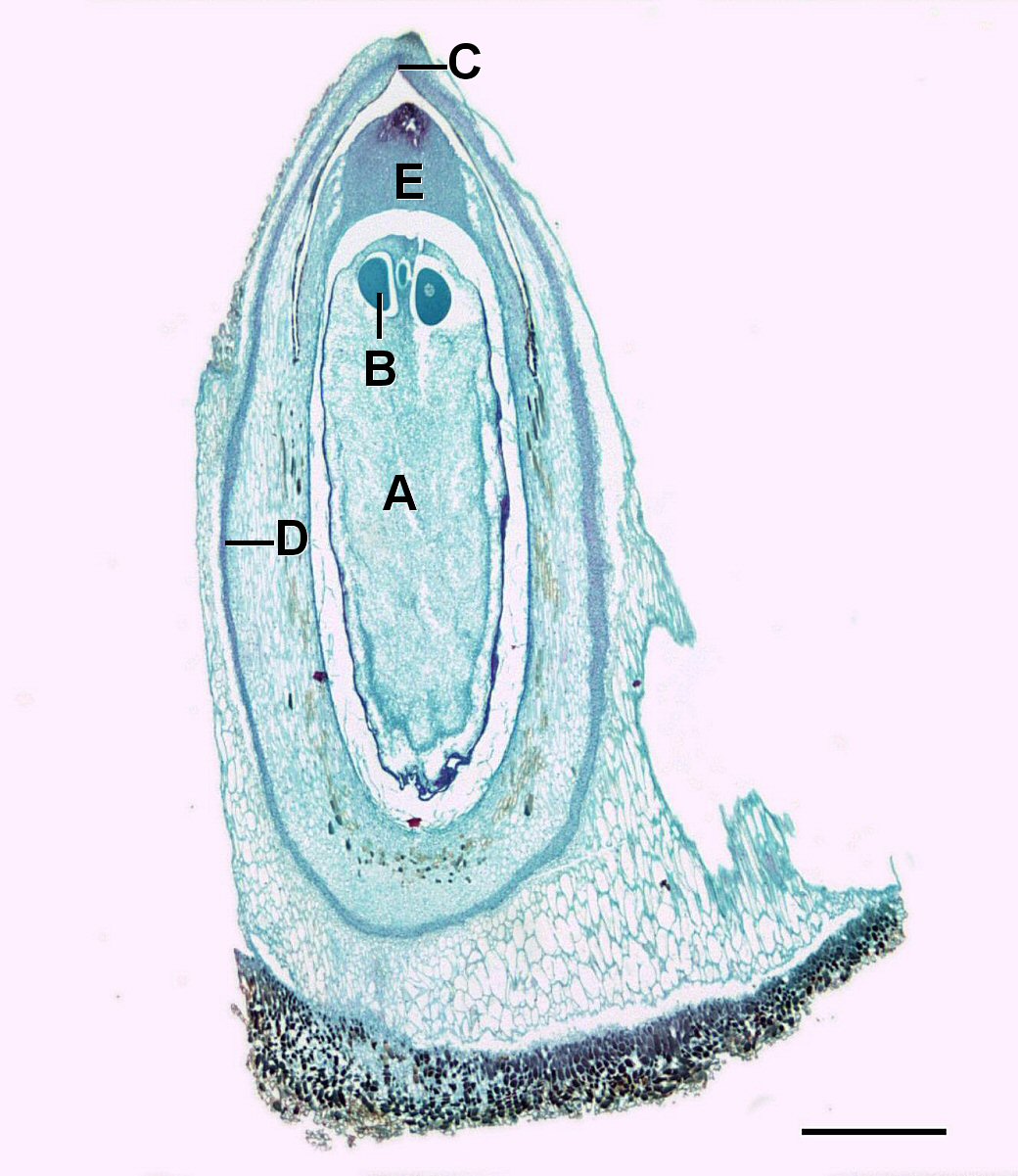
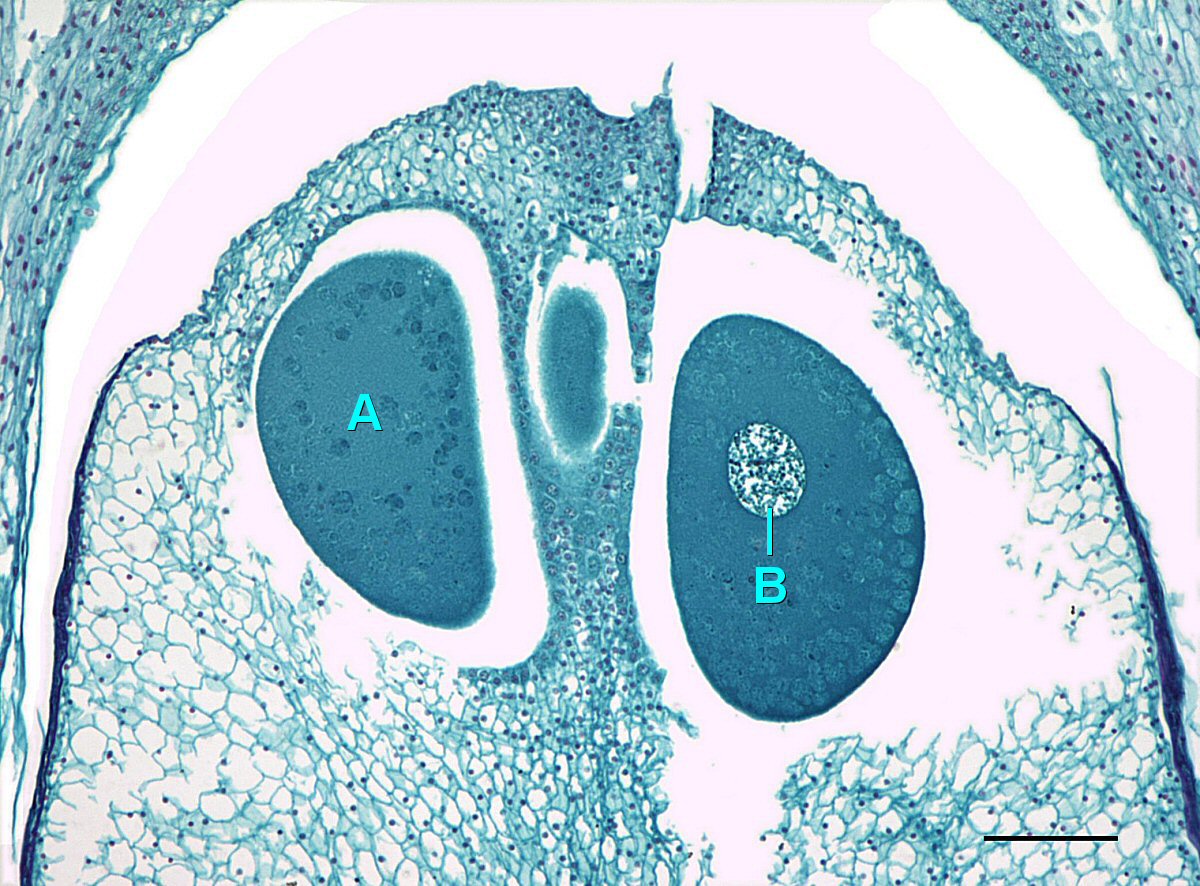
Un grano de polen se transportará al viento y, si tiene suerte, aterrizará sobre un cono de semilla. El cono de semilla tiene una gota de líquido azucarado que secreta, luego se retrae, tirando del polen hacia el óvulo. Esto estimula a la célula tubular a germinar un tubo de polen, mientras que la célula generativa se divide por mitosis para producir dos espermatozoides. Estos espermatozoides viajan por el tubo de polen, a través del micropilo, y hacia un archegonio donde se fertilizará un óvulo. Cuando ocurre la fertilización, el micropilo se cierra y el tegumento se convierte en la capa de la semilla.


El cigoto crecerá y se desarrollará como un embrión, nutrido por el tejido megagametofítico, así como el nucello. Si miras en una sección larga de una semilla de pino, puedes ver la RAM y SAM del embrión. La semilla será dispersada por el viento o los animales y germinará para crecer una vez más hasta convertirse en un pino diploide.
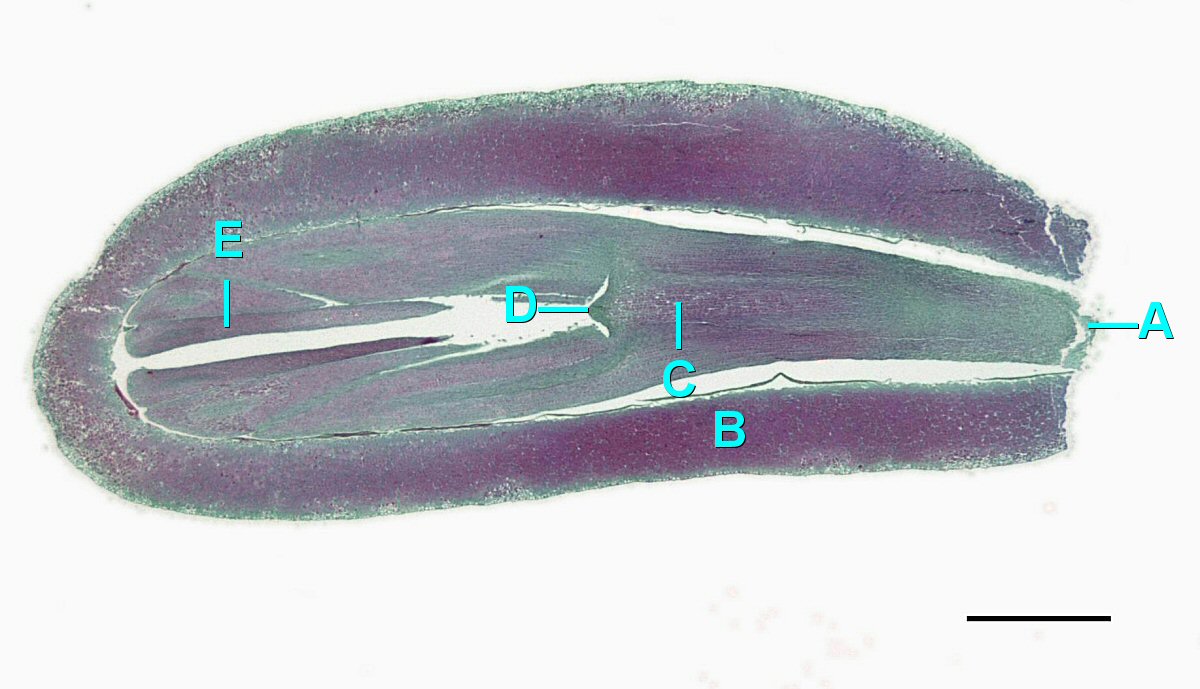
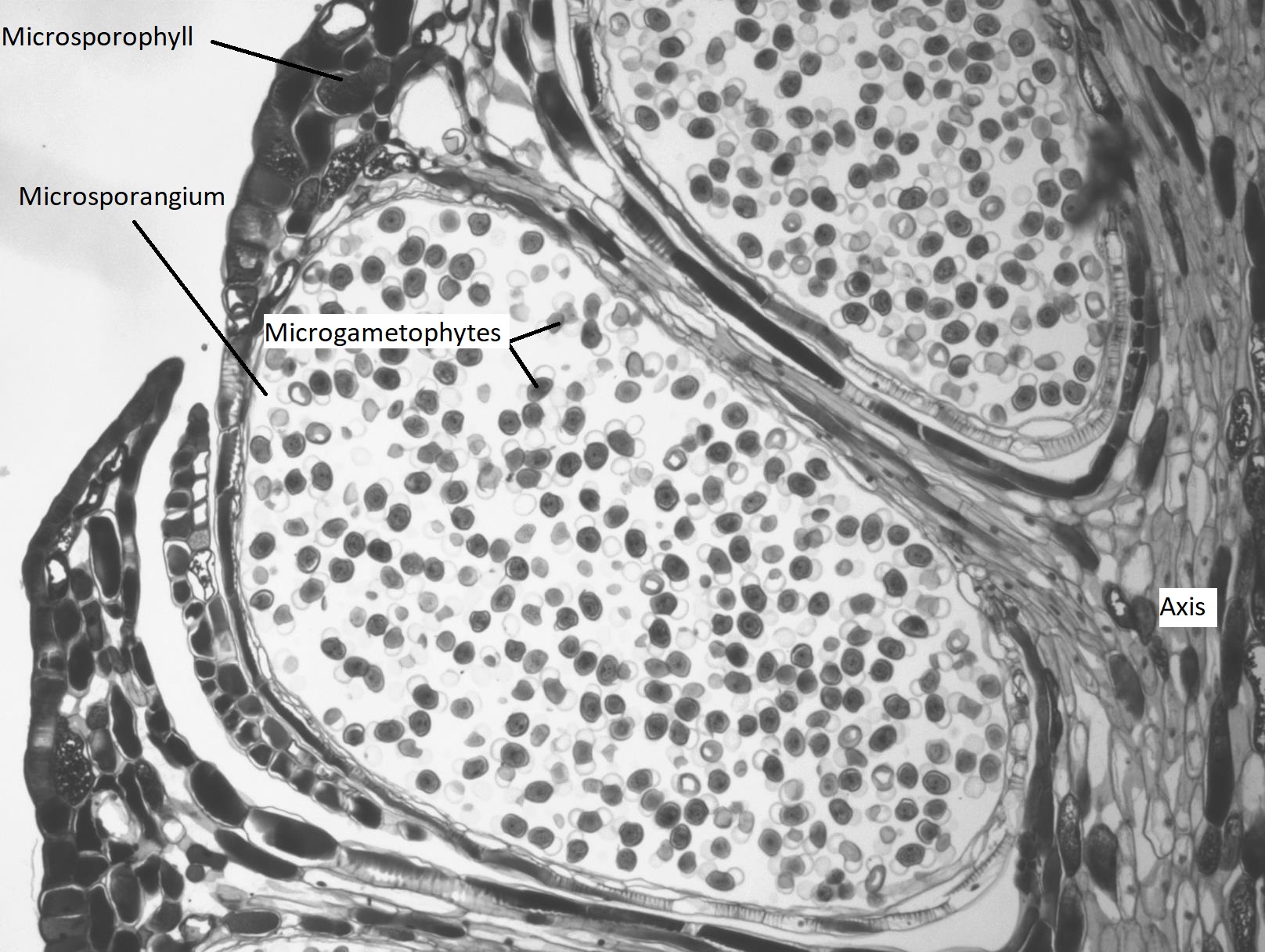
Conos de Polen

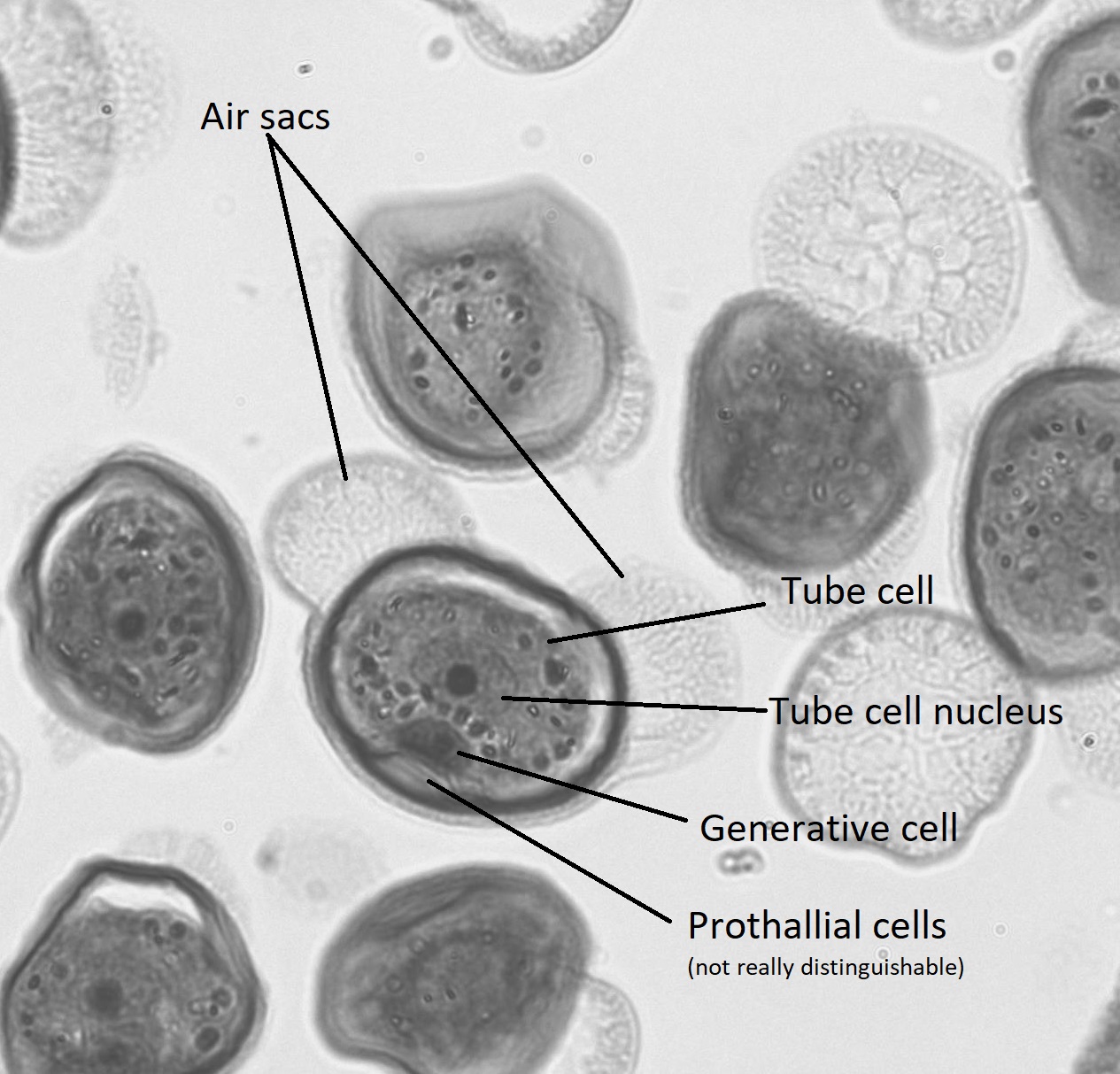
El microgametofito en las gimnospermas es el grano de polen “alado” de cuatro células. Dentro del grano de polen, se puede distinguir la célula generativa y el núcleo de la célula tubular. Las dos células protaliales no son aparentes bajo el microscopio. A ambos lados del grano de polen, emergen dos estructuras similares a orejas. Estos sacos de aire pueden ayudar a orientar el grano de polen hacia el óvulo.
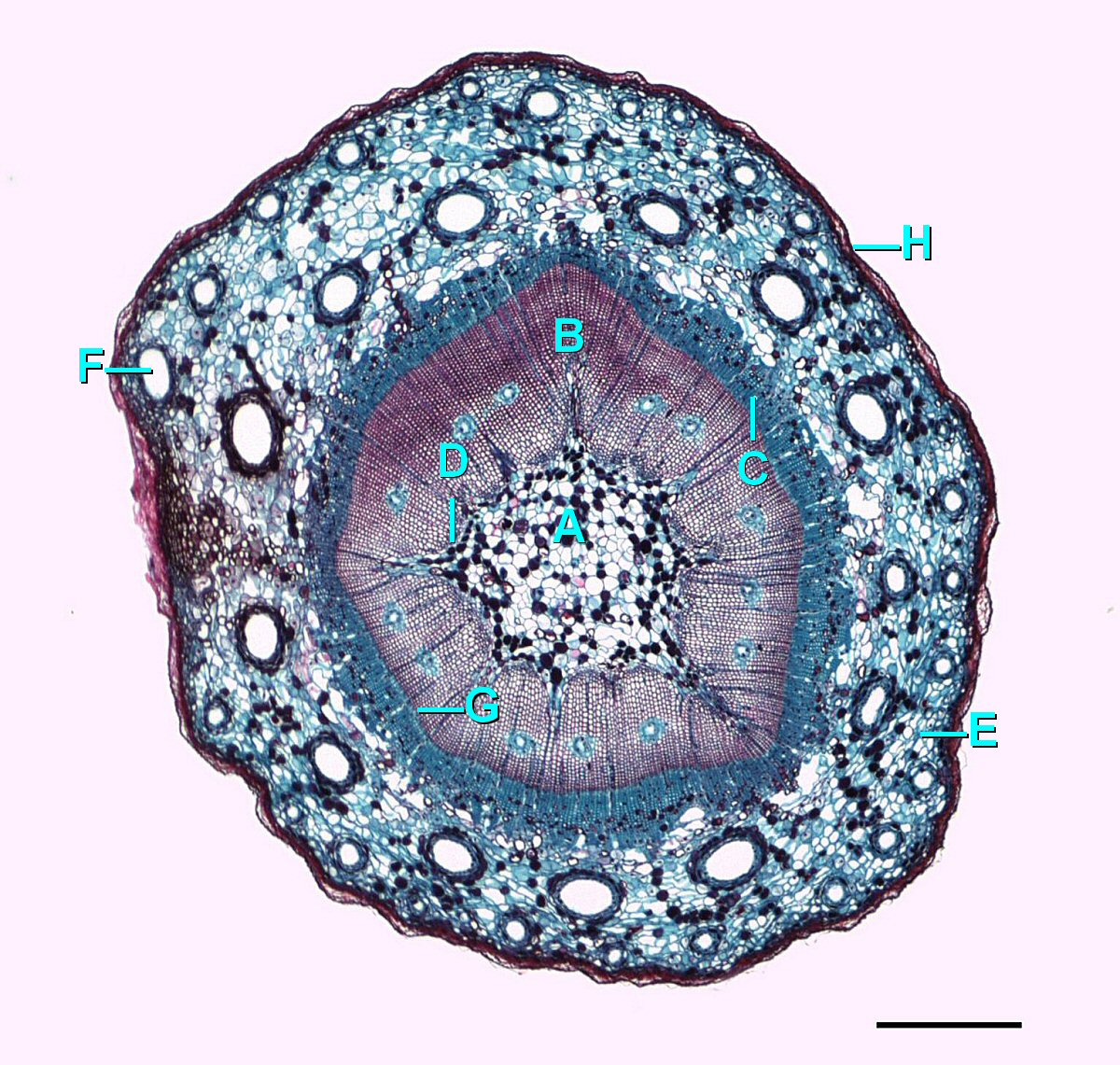
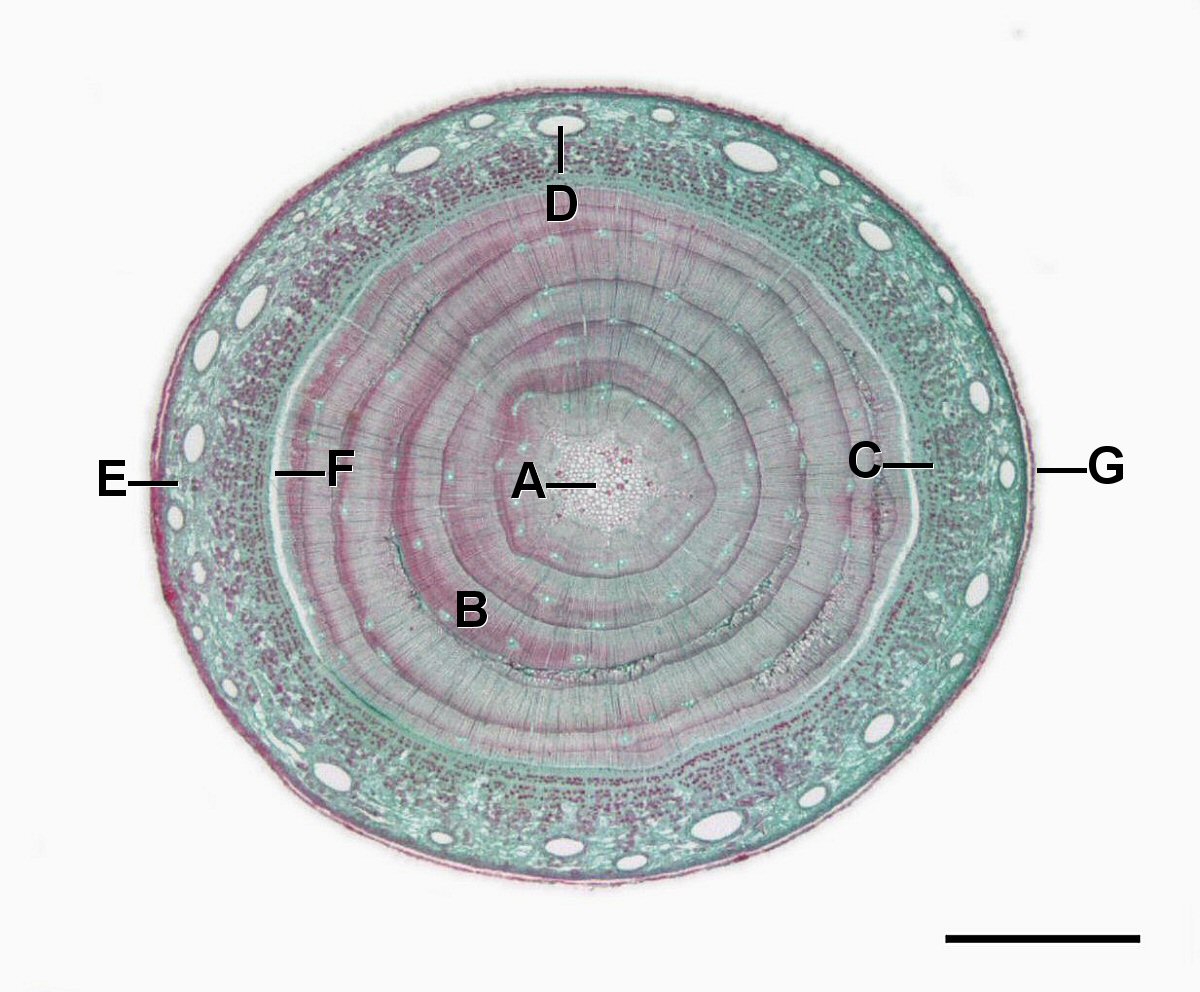
Crecimiento Secundario
_LI.jpg)
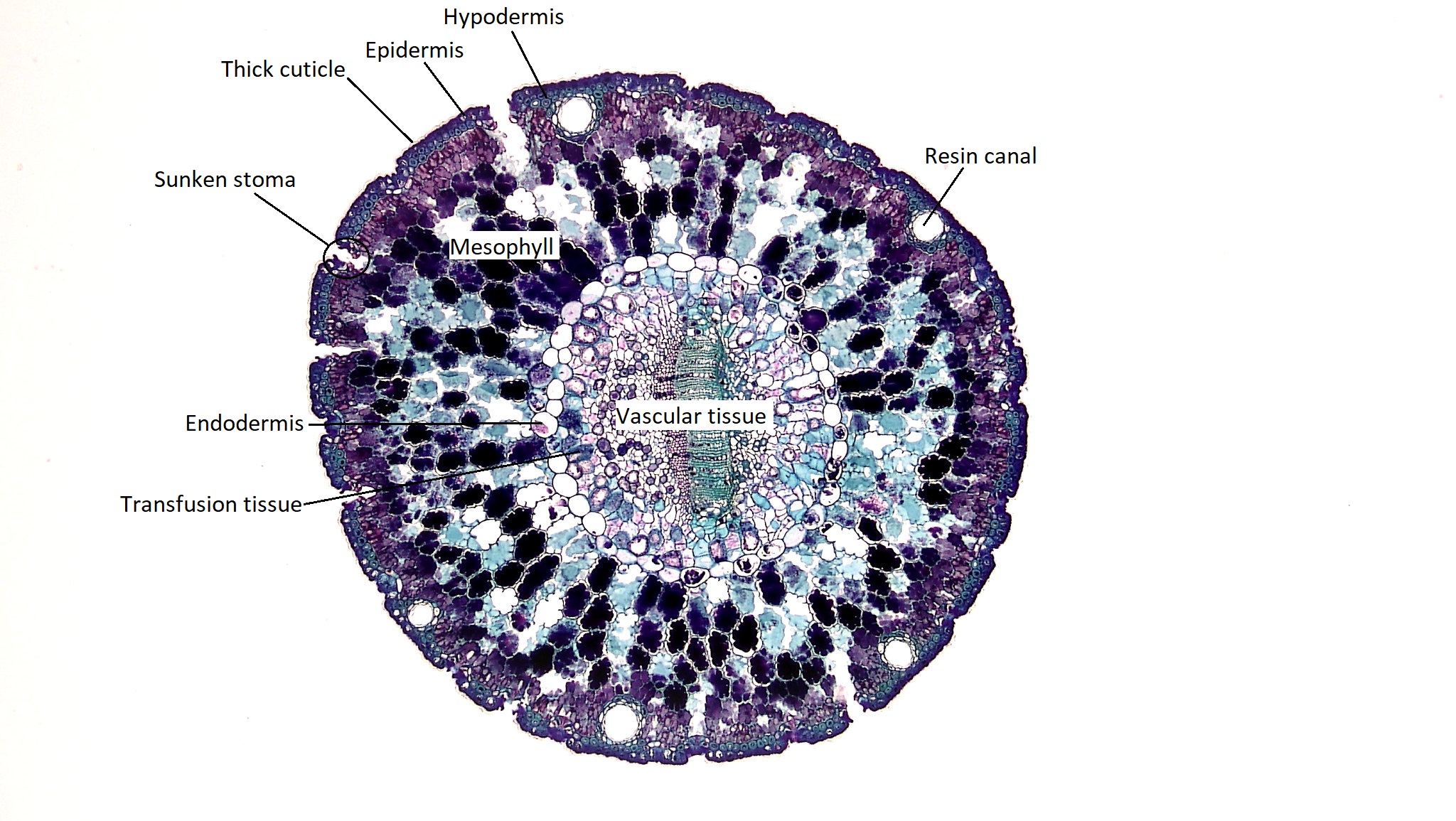
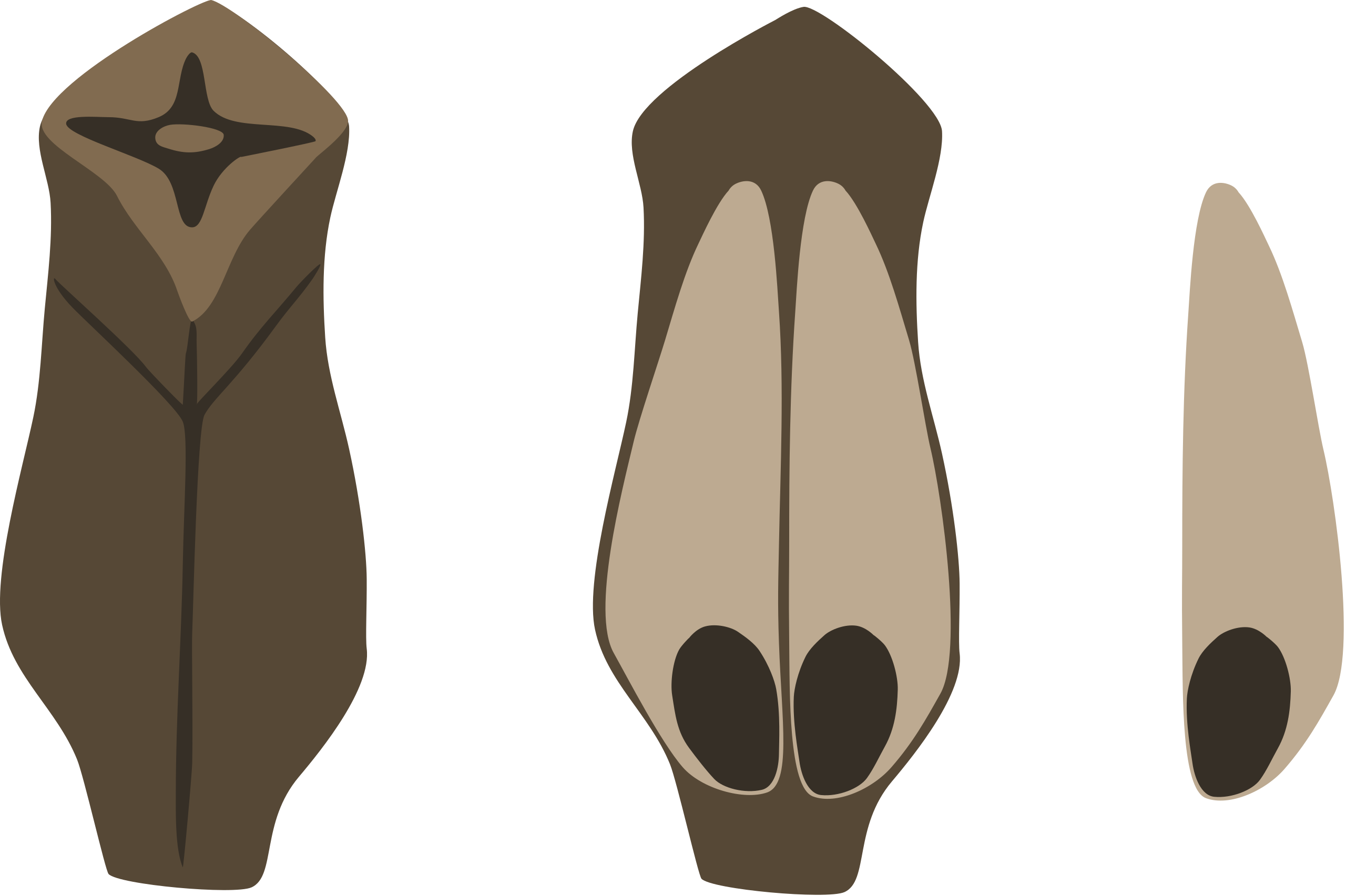
Hojas Xerofíticas
Las hojas xerofíticas están adaptadas para soportar condiciones de sequía. En las coníferas, vemos una amplia gama de hojas xerofíticas con diferentes morfologías que pueden ser moldeadas por su entorno local. Considera las hojas de la secoya costera y la secuoya gigante, que se muestran a continuación. Aunque estos dos árboles pertenecen a diferentes géneros, Sequoia y Sequoiadendron, respectivamente, son taxones hermanos. Sin embargo, la secoya costera se ha adaptado a la vida en la costa, donde la secuoya gigante ha evolucionado en bosques interiores de mayor elevación con condiciones climáticas mucho más extremas. ¿Cómo se puede ver esto en la estructura de sus hojas?