
1.5: La ley de la segregación




- Última actualización
- 29 oct 2022
- Guardar como PDF
( \newcommand{\kernel}{\mathrm{null}\,}\)
Puntos clave:
- Gregor Mendel estudió la herencia de rasgos en plantas de guisante. Propuso un modelo donde pares de “elementos heredables”, o genes, especificaban rasgos.
- Los genes vienen en diferentes versiones, o alelos. Un alelo dominante esconde un alelo recesivo y determina la apariencia del organismo.
- Cuando un organismo fabrica gametos, cada gameto recibe solo una copia génica, la cual se selecciona aleatoriamente. Esto se conoce como la ley de la segregación.
- Un cuadrado Punnett se puede utilizar para predecir genotipos (combinaciones de alelos) y fenotipos (rasgos observables) de crías a partir de cruces genéticos.
- Se puede utilizar un cruce de prueba para determinar si un organismo con un fenotipo dominante es homocigoto o heterocigótico.
Introducción
Hoy en día, sabemos que muchas de las características de las personas, desde el color del cabello hasta la altura y el riesgo de diabetes, están influenciadas por los genes. También sabemos que los genes son la forma en que los padres transmiten características a sus hijos (incluyendo cosas como hoyuelos, o —en el caso de mi padre y yo— una voz terrible para cantar). En los últimos cien años, hemos llegado a entender que los genes son en realidad trozos de ADN que se encuentran en los cromosomas y especifican proteínas.
Pero, ¿siempre sabíamos esas cosas? ¡No por un tiro lejano! Hace unos 150 años, un monje llamado Gregor Mendel publicó un artículo que propuso por primera vez la existencia de genes y presentó un modelo de cómo fueron heredados. El trabajo de Mendel fue el primer paso en un largo camino, involucrando a muchos científicos trabajadores, eso nos ha llevado a nuestra comprensión actual de los genes y lo que hacen.
En este artículo, rastrearemos los experimentos y razonamientos que llevaron a Mendel a formular su modelo para la herencia de genes individuales.
El modelo de Mendel: Comenzó con una relación 3:1
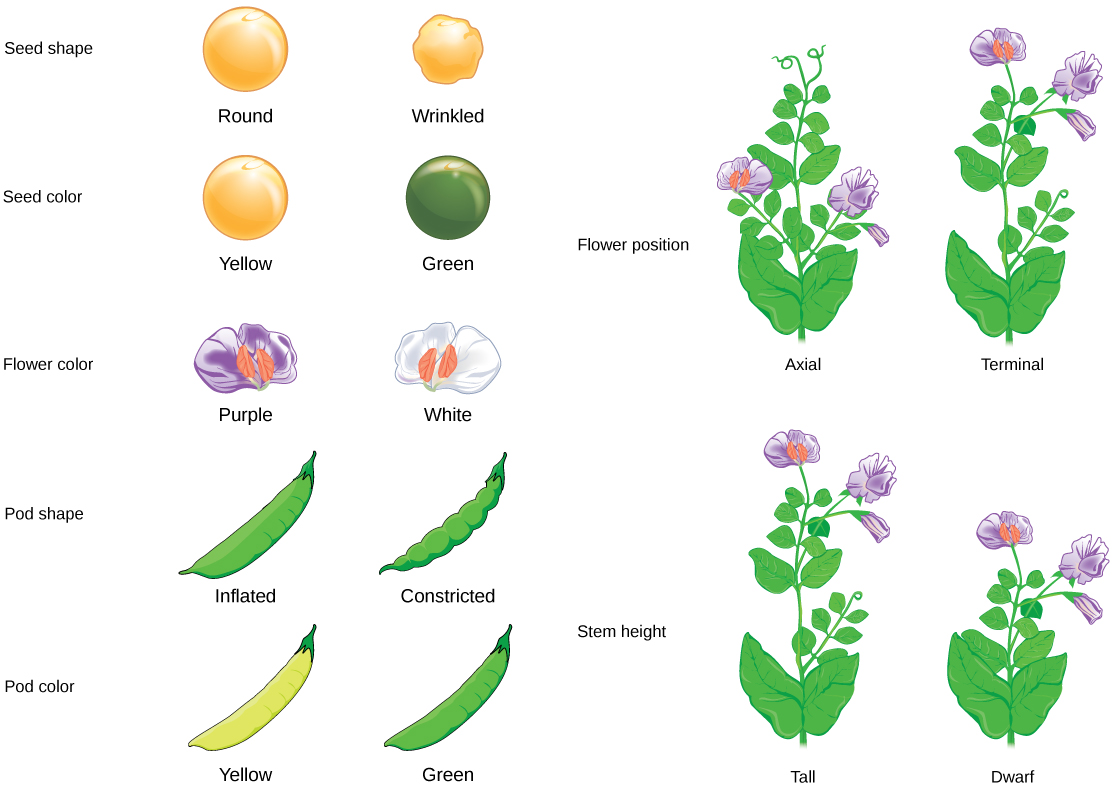
Mendel estudió la genética de las plantas de guisante, y trazó la herencia de una variedad de características, incluyendo el color de la flor, la posición de la flor, el color de la semilla y la forma de la semilla. Para ello, comenzó cruzando plantas parentales de pura reproducción con distintas formas de característica, como las flores violetas y blancas. La cría pura solo significa que la planta siempre hará más descendencia como ella, cuando se autofertiliza a lo largo de muchas generaciones.
- [¿Qué es la autofecundación?]
-
En la autofecundación, espermatozoides y óvulos de la misma planta de guisante se combinan dentro de una flor cerrada, produciendo semillas con una sola planta como madre y padre.
¿Qué resultados encontró Mendel en sus cruces para el color de la flor? En la generación parental, o P, Mendel cruzó una planta de flores violetas de reproducción pura a una planta de flores blancas de reproducción pura. Cuando recolectó y sembró las semillas producidas en este cruce, Mendel encontró que el 100 por ciento de las plantas de la siguiente generación, o la generación F 1, tenían flores violetas.
La sabiduría convencional en ese momento habría predicho que las flores híbridas deberían ser violetas pálidas, es decir, que los rasgos de los padres deberían mezclarse en la descendencia. En cambio, los resultados de Mendel mostraron que el rasgo de flor blanca había desaparecido por completo. Llamó al rasgo que era visible en la generación F 1 (flores violetas) el rasgo dominante, y el rasgo que estaba oculto o perdido (flores blancas) el rasgo recesivo.
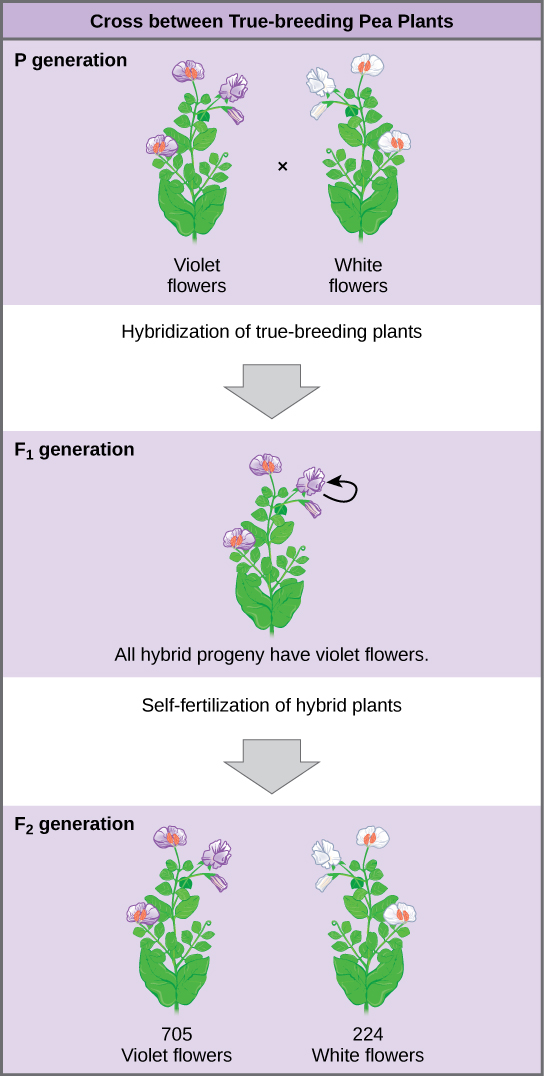
Es importante destacar que Mendel no detuvo allí su experimentación. En cambio, dejó que las plantas F 1 se autofertilizaran. Entre sus crías, llamada la generación F 2, encontró que 705 plantas tenían flores violetas y 224 flores blancas. Esta fue una relación de 3.15 flores violetas a una flor blanca, o aproximadamente 3:1.
Esta relación 3:1 no fue una platija. Para las otras seis características que Mendel examinó, tanto las generaciones F 1 como F 2 se comportaron de la misma manera que lo hicieron para el color de la flor. Uno de los dos rasgos desaparecería completamente de la generación F 1, solo para reaparecer en la generación F 2 en una proporción de aproximadamente 3:1.
- [Ver los datos de Mendel para las siete características]
-
Carácter Rasgo dominante Rasgo recesivo Generación F 2 Relación F 2 Color de semilla Amarillo Verde 6022 amarillo, 2001 verde 3. 01:1 Forma de semilla Redonda Arrugada 5474 redondo, 1850 arrugado 2. 96:1 Color de la vaina Verde Amarillo 428 verde, 152 amarillo 2. 82:1 Forma de vaina Inflado Constreñido 882 inflado, 299 constreñido 2. 95:1 Altura de la planta Tall Corto (enano) 787 alto, 277 corto 2. 84:1 Color de la flor Morado Blanco 705 púrpura, 224 blanco 3. 15:1 Posición de la flor Axial (a lo largo de rama Terminal (final de rama) 651 axial, 207 terminal 3. 14:1 Adaptado de "Los resultados de las hibridaciones de guisantes de jardín de Mendel”, de OpenStax College, Biology (CC BY 3.0).
Al final resultó que, la proporción 3:1 era una pista crucial que dejó a Mendel romper el rompecabezas de la herencia. Echemos un vistazo más de cerca a lo que descubrió Mendel.
El modelo de herencia de Mendel
A partir de sus resultados (incluyendo esa proporción mágica 3:1), Mendel ideó un modelo para la herencia de características individuales, como el color de la flor.
En el modelo de Mendel, los padres pasan “factores heredables”, que ahora llamamos genes, que determinan los rasgos de la descendencia. Cada individuo tiene dos copias de un gen dado, como el gen para el color de la semilla (gen Y) que se muestra a continuación. Si estas copias representan diferentes versiones, o alelos, del gen, un alelo, el dominante, puede ocultar al otro alelo, el recesivo. Para el color de la semilla, el alelo amarillo dominante Y oculta el alelo verde recesivo y.
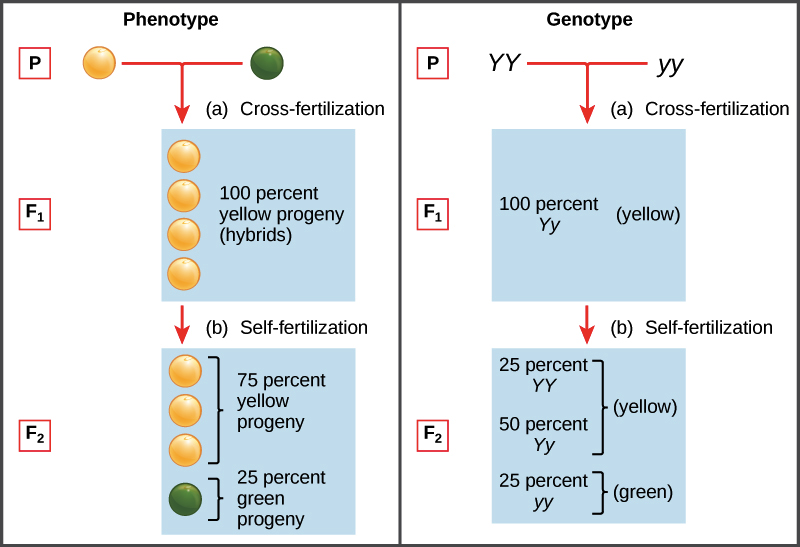
El conjunto de alelos portados por un organismo se conoce como su genotipo. El genotipo determina el fenotipo, características observables de un organismo. Cuando un organismo tiene dos copias del mismo alelo (digamos, YY o yy), se dice que es homocigótico para ese gen. Si, en cambio, tiene dos copias distintas (como Yy), podemos decir que es heterocigótico. El fenotipo también puede verse afectado por el ambiente en muchos casos de la vida real, aunque esto no tuvo un impacto en el trabajo de Mendel.
El modelo de Mendel: La ley de la segregación
Hasta el momento, tan bien. Pero este modelo por sí solo no explica por qué Mendel vio los patrones exactos de herencia que hizo. En particular, no da cuenta de la relación 3:1. Para ello, necesitamos la ley de segregación de Mendel.
De acuerdo con la ley de segregación, sólo una de las dos copias génicas presentes en un organismo se distribuye a cada gameto (óvulo o espermatozoide) que realiza, y la asignación de las copias génicas es aleatoria. Cuando un óvulo y un espermatozoide se unen en la fertilización, forman un nuevo organismo, cuyo genotipo consiste en los alelos contenidos en los gametos. El siguiente diagrama ilustra esta idea:
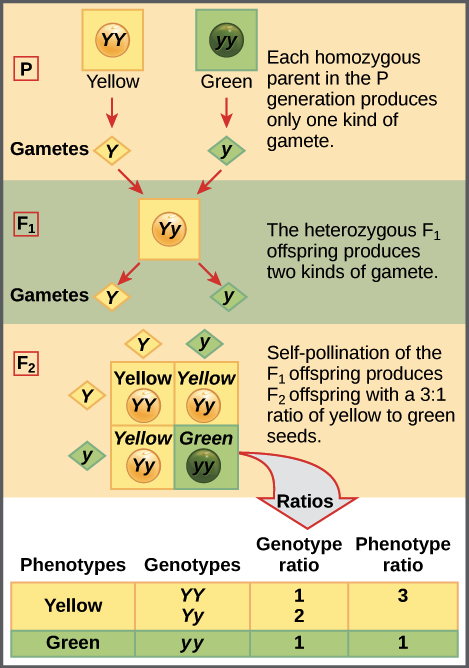
La caja de cuatro cuadrados que se muestra para la generación F 2 se conoce como un cuadrado Punnett. Para preparar un cuadrado Punnett, todos los gametos posibles hechos por los padres se escriben a lo largo de la parte superior (para el padre) y lateral (para la madre) de una cuadrícula. Aquí, al ser autofecundación, la misma planta es a la vez madre y padre.
Luego se hacen las combinaciones de óvulo y esperma en las cajas de la tabla, lo que representa la fertilización para hacer nuevos individuos. Debido a que cada cuadrado representa un evento igualmente probable, podemos determinar las proporciones de genotipo y fenotipo contando los cuadrados.
- [¿Por qué todas las cajas son igualmente probables?]
-
Un punto clave de la ley de segregación es que las dos copias genéticas de un progenitor se distribuyen aleatoriamente a sus gametos. Así, para un heterocigoto Yy, es igualmente probable que se hagan gametos Y e y: 50% de los espermatozoides y óvulos tendrán un alelo Y, 50% tendrán un alelo y, y lo mismo será cierto para los óvulos.
Dado que cada tipo de gameto es igualmente común, cada evento de fertilización (encuentro de gametos, correspondiente a un cuadrado de la tabla) también tiene las mismas posibilidades de suceder. Así, las casillas de la tabla representan cuatro eventos de igual probabilidad.
Dado que la tabla contiene 1 caja con un genotipo YY, 2 cajas con un genotipo Yy y 1 caja con un genotipo yy, esperaríamos ver plantas YY, Yy y yy en una proporción de 1:2:1 en la generación F 2. Dado que tanto las plantas YY como Yy son amarillas, esta relación de genotipos se traduce en una relación de fenotipos de 3:1 de plantas sembradas de amarillo a verde, casi exactamente lo que observó Mendel.
Las plantas YY y yy aparecen cada una en un solo cuadrado, mientras que las plantas Yy se encuentran en dos cuadrados. Esto se debe a que hay dos eventos diferentes de fertilización que conducen a una planta Yy: la fusión de un óvulo Y y un espermatozoide y, o la fusión de un óvulo y y un espermatozoide Y. Cualquiera de los dos eventos es igualmente probable y tiene la misma probabilidad que un evento de fertilización YY o yy.
El cruce de prueba
A Mendel también se le ocurrió una manera de averiguar si un organismo con un fenotipo dominante (como una planta de guisante de siembra amarilla) era un heterocigoto (Yy) o un homocigoto (YY). Esta técnica se llama cruce de prueba y todavía es utilizada por los criadores de plantas y animales en la actualidad.
En un cruce de prueba, el organismo con el fenotipo dominante se cruza con un organismo homocigótico recesivo (p. ej., de siembra verde):
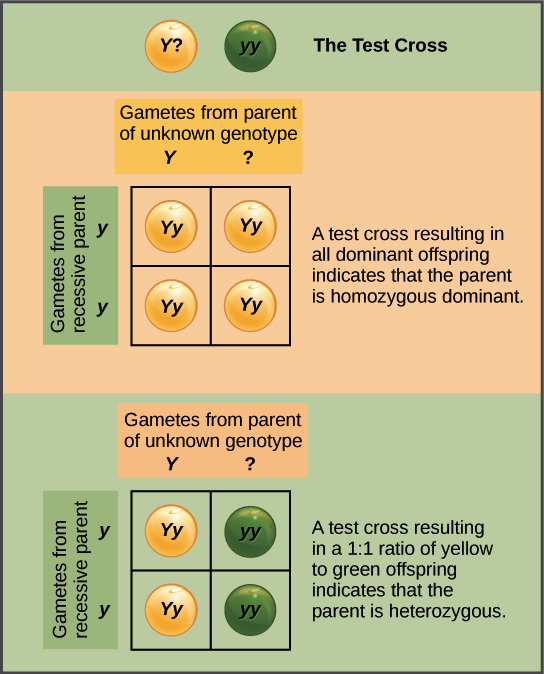
Si el organismo con el fenotipo dominante es homocigótico, entonces toda la descendencia F 1 obtendrá un alelo dominante de ese progenitor, será heterocigoto y mostrará el fenotipo dominante. Si el organismo con el organismo fenotipo dominante es en cambio un heterocigoto, la descendencia F 1 será mitad heterocigotos (fenotipo dominante) y mitad homocigotos recesivos (fenotipo recesivo).
El hecho de que obtengamos una proporción 1:1 en este segundo caso es otra confirmación de la ley de segregación de Mendel.
¿Ese es el modelo completo de herencia de Mendel?
¡No del todo! Hemos visto todo el modelo de Mendel para la herencia de genes individuales. Sin embargo, el modelo completo de Mendel también abordó si los genes para diferentes características (como el color de la flor y la forma de la semilla) influyen en la herencia de los demás. Puedes conocer más sobre el modelo de Mendel para la herencia de múltiples genes en el artículo de ley del surtido independiente.
Una cosa que me parece bastante sorprendente es que Mendel pudo averiguar todo su modelo de herencia simplemente a partir de sus observaciones de las plantas de guisante. Esto no fue porque fuera una especie de súper genio loco, sino más bien, porque era muy cuidadoso, persistente y curioso, y también porque pensaba matemáticamente en sus resultados (por ejemplo, la relación 3:1). Estas son algunas de las cualidades de un gran científico, ¡unas que cualquiera, en cualquier lugar, puede desarrollar!
Comprueba tu comprensión
Consulta1.5.1
- [Pista]
-
Cuando se cruzan dos organismos de raza pura con diferentes formas de una característica, la descendencia de la cruz puede mostrar solo una forma de la característica, mientras que la otra forma puede estar oculta.
El rasgo que es visible en la descendencia de la cruz se llama rasgo dominante, mientras que el rasgo que se oculta se llama rasgo recesivo.
En este ejemplo, el pelaje bronceado es el rasgo dominante, porque es visible en la descendencia de la cruz. El pelaje negro es el rasgo recesivo, porque está oculto en la descendencia de la cruz.
El pelaje bronceado es dominante, mientras que el negro es recesivo.
Consulta1.5.2
- [Pista]
-
Genotipo, el conjunto de alelos que lleva un organismo, determina el fenotipo, su aspecto exterior o características. Sin embargo, el ambiente influye en cómo el genotipo se traduce en fenotipo, por lo que el fenotipo de un organismo es típicamente el producto tanto del genotipo como del ambiente.
El genotipo determina el fenotipo, pero con influencia del ambiente.
Colaboradores y Atribuciones
- [Atribución y referencias]
-
Atribución:
Este artículo es un derivado modificado de los siguientes artículos:
- “Los experimentos de Mendel”, en Principios de Biología, de Robert Bear, David Rintoul, Bruce Snyder, Martha Smith-Caldas, Christopher Herren y Eva Horne, OpenStax, CC BY 4.0. Descarga gratis el artículo original en http://cnx.org/contents/db89c8f8-a27c-4685-ad2a-19d11a2a7e2e@24.18.
- “Leyes de herencia”, en Principios de Biología, de Robert Bear, David Rintoul, Bruce Snyder, Martha Smith-Caldas, Christopher Herren, y Eva Horne, OpenStax, CC BY 4.0. Descarga gratis el artículo original en http://cnx.org/contents/db89c8f8-a27c-4685-ad2a-19d11a2a7e2e@24.18.
- “Los experimentos de Mendel y las leyes de la probabilidad”, por OpenStax College, Biology (CC BY 3.0). Descarga gratis el artículo original en http://cnx.org/contents/185cbf87-c72e-48f5-b51e-f14f21b5eabd@9.85.
El artículo modificado está licenciado bajo una licencia CC BY-NC-SA 4.0.
Referencias adicionales:
Ding, Z. (2009). Color de ojos. En Stanford en el Tech: Entendiendo la genética. Recuperado de http://genetics.thetech.org/ask/ask316.
Miko, I. (2008). Gregor Mendel y los principios de herencia. Educación de la Naturaleza, 1 1 (1) :134. Recuperado de http://www.nature.com/scitable/topicpage/gregor-mendel-and-the-principles-of-inheritance-593.
Cruce monohíbrido. (2015, 18 de septiembre). Recuperado el 14 de noviembre del 2015 de Wikipedia: https://en.wikipedia.org/wiki/Monohybrid_cross.
Fenotipo. (2015, 17 de octubre). Recuperado el 14 de noviembre del 2015 de Wikipedia: https://en.wikipedia.org/wiki/Phenotype.
Punnet. (2015, 9 de agosto). Recuperado el 18 de noviembre del 2015 de Wikipedia: https://en.wikipedia.org/wiki/Punnet.
Plaza Punnett. (2015, 8 de noviembre). Recuperado el 14 de noviembre del 2015 de Wikipedia: https://en.wikipedia.org/wiki/Punnett_square.
Purves, W. K., Sadava, D. E., Orians, G. H., y Heller, H.C. (2004). Genética: Mendel y más allá. En la vida: La ciencia de la biología (7ª ed., pp. 187-212). Sunderland, MA: Asociados Sinauer.
Raven, P. H., Johnson, G. B., Mason, K. A., Losos, J. B., y Singer, S. R. (2014). Patrones de herencia. En Biología (10a ed., AP ed., pp. 221-238). Nueva York, NY: McGraw-Hill.
Reece, J. B., Urry, L. A., Caín, M. L., Wasserman, S. A., Minorsky, P. V., y Jackson, R. B. (2011). Mendel y la idea genética. En biología Campbell (10a ed., pp. 267-291). San Francisco, CA: Pearson.
Starr, B. (2006, 20 de diciembre). Color de ojos [respuesta]. En Stanford en el Tech: Entendiendo la genética. Recuperado a partir de http://genetics.thetech.org/ask/ask203
Strehlow, A. T. (2005, 9 de marzo). Otros rasgos [respuesta]. En Stanford en el Tech: Entendiendo la genética. Recuperado de http://genetics.thetech.org/ask/ask98.
White, D. y Rabago-Smith, M. (2011). Asociaciones genotipo-fenotipo y color de ojos humanos. Revista de Genética Humana, 56, 5-7. http://dx.doi.org/10.1038/jhg.2010.126.