
3.2: Herencia ligada al X
- Page ID
- 57524
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Puntos clave:
- En humanos y otros mamíferos, el sexo biológico está determinado por un par de cromosomas sexuales: XY en machos y XX en hembras.
- Se dice que los genes en el cromosoma X están ligados a X. Los genes ligados al X tienen patrones de herencia distintivos porque están presentes en diferentes números en hembras (XX) y machos (XY).
- Los trastornos genéticos humanos ligados al X son mucho más comunes en hombres que en mujeres debido al patrón de herencia ligada al cromosoma X.
Introducción
Si eres un ser humano (¡lo cual parece una buena apuesta!) , la mayoría de tus cromosomas vienen en pares homólogos. Los dos cromosomas de un par homólogo contienen la misma información básica —es decir, los mismos genes en el mismo orden— pero pueden portar diferentes versiones de esos genes.
¿Todos tus cromosomas están organizados en pares homólogos? La respuesta depende de si eres (cromosómicamente) masculino.
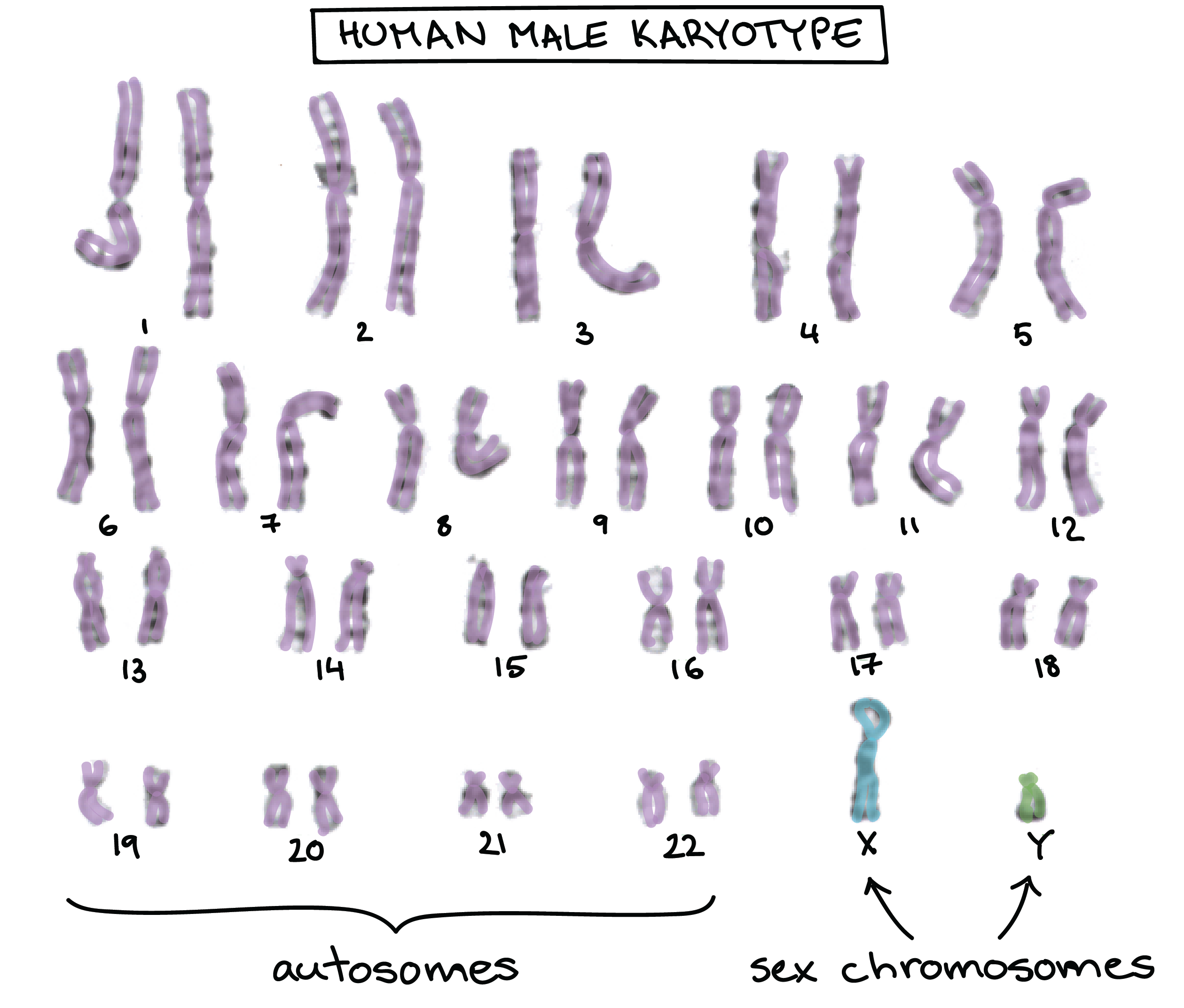
- Un macho humano tiene dos cromosomas sexuales, el X y el Y. A diferencia de los 44 autosomas (cromosomas no sexuales), los X e Y no portan los mismos genes y no se consideran homólogos.
- En lugar de una X y una Y, una hembra humana tiene dos cromosomas X. Estos cromosomas X forman un par homólogo de buena fe.
Debido a que los cromosomas sexuales no siempre vienen en pares homólogos, los genes que portan muestran patrones de herencia únicos y distintivos.
Cromosomas sexuales en humanos
Los cromosomas X e Y humanos determinan el sexo biológico de una persona, con XX especificando hembra y XY especificando masculino. Aunque el cromosoma Y contiene una pequeña región de similitud con el cromosoma X para que puedan emparejarse durante la meiosis, el cromosoma Y es mucho más corto y contiene muchos menos genes.
Para ponerle algunos números, el cromosoma X tiene alrededor de 800-900 genes codificadores de proteínas con una amplia variedad de funciones, mientras que el cromosoma Y tiene apenas 60-70 genes codificadores de proteínas, aproximadamente la mitad de los cuales están activos solo en los testículos (órganos productores de espermatozoides) 1,2,3,4.
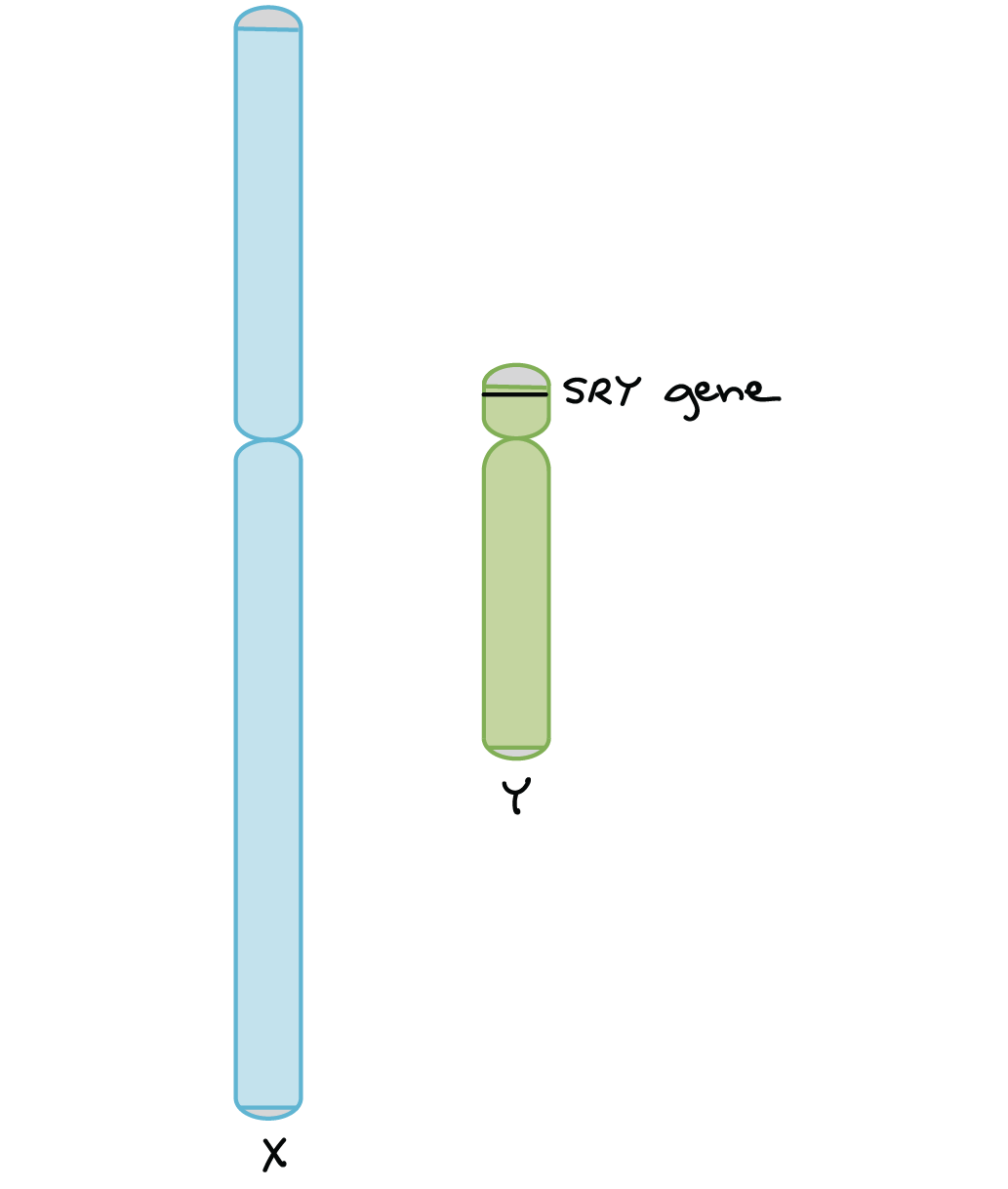
El cromosoma Y humano juega un papel clave en la determinación del sexo de un embrión en desarrollo. Esto se debe principalmente a un gen llamado SRY (“región determinante del sexo de Y”). SRY se encuentra en el cromosoma Y y codifica una proteína que activa otros genes requeridos para el desarrollo masculino 5,6.
- XX embriones no tienen SRY, por lo que se desarrollan como hembras.
- Los embriones XY sí tienen SRY, por lo que se desarrollan como machos.
En casos raros, los errores durante la meiosis pueden transferir SRY del cromosoma Y al cromosoma X. Si un cromosoma X portador de SRY fertiliza un óvulo normal, producirá un embrión cromosómicamente femenino (XX) que se desarrolla como macho 7. Si un cromosoma Y deficiente en SRY fertiliza un óvulo normal, producirá un embrión cromosómicamente masculino (XY) que se desarrolla como hembra 8.
- [¿Todas las especies con cromosomas X e Y tienen el gen SRY?]
-
No, no todos ellos. Los mecanismos moleculares de determinación del sexo varían mucho entre especies, incluso aquellos que utilizan un sistema de determinación del sexo X-Y. En la mayoría de los mamíferos placentarios, el SRY se encuentra en el cromosoma Y y se utiliza para la determinación del sexo. Sin embargo, el gen SRY no está presente en otras especies con sistemas de determinación del sexo X-Y, como moscas de la fruta y otros insectos 9.
Los cromosomas X e Y han evolucionado independientemente muchas veces
Para entender cómo esto es posible, es útil tener en cuenta que “X” e “Y” son solo etiquetas genéricas aplicadas a los cromosomas dimórficos (di - = dos, - morfo = forma), o disímiles, que se encuentran en especies con sistemas de determinación del sexo X-Y 10. X es cualquier cromosoma del que la hembra tenga dos, mientras que Y es el cromosoma emparejado con una sola X en el macho. La naturaleza de los cromosomas —qué genes hay en ellos y cómo determinan la masculinidad o la feminidad— puede ser bastante diferente entre especies.
Los cromosomas sexuales han evolucionado independientemente muchas veces 11. Así, aunque las moscas de la fruta y los humanos tienen ambos cromosomas X e Y que coinciden con la definición anterior, y aunque las combinaciones de cromosomas XY y XX corresponden a masculinidad y femalidad, respectivamente, en ambas especies, el mecanismo de determinación del sexo en las moscas es muy diferente al de los humanos.
Ejemplo: Humanos vs moscas de la fruta
En humanos, como se describió anteriormente, el cromosoma Y especifica la masculinidad a través de la acción del gen SRY, un regulador maestro del desarrollo masculino.
En las moscas, la presencia de un solo cromosoma X especifica masculinidad, mientras que la presencia de dos Xs especifica femalidad, independientemente de la presencia o ausencia de un cromosoma Y. (Más específicamente, parece que el sexo está determinado por la relación de cromosomas X a autosomas, con una relación 1:2 especificando masculinidad y una relación 2:2 especificando femalidad. Estas proporciones se “miden” a través de los niveles de proteínas producidas por genes específicos en el cromosoma X y en los autosomas). 12
Debido a estas diferencias en los mecanismos de determinación del sexo, una mosca XXY se desarrollará como hembra (ver artículo sobre los experimentos de Thomas Hunt Morgan), mientras que un humano XXY se desarrollará como macho (en una condición conocida como síndrome de Klinefelter; ver artículo sobre cambios cromosómicos a gran escala).
Genes ligados al X
Cuando un gen está presente en el cromosoma X, pero no en el cromosoma Y, se dice que está ligado a X. Los genes ligados al X tienen diferentes patrones de herencia que los genes en cromosomas no sexuales (autosomas). Eso se debe a que estos genes están presentes en diferentes números de copias en machos y hembras.
- [¿Qué pasa con los genes en la Y?]
-
Los genes en la X o en la Y tienen patrones de herencia inusuales y se llaman ligados al sexo, pero el enlace X es mucho más común que el enlace Y (ya que hay muchos más genes en la X que en la Y).
Dado que una hembra tiene dos cromosomas X, tendrá dos copias de cada gen ligado al X. Por ejemplo, en la mosca de la fruta Drosophila (que, al igual que los humanos, tiene XX hembras y machos XY), hay un gen de color de ojos llamado blanco que se encuentra en el cromosoma X, y una mosca hembra tendrá dos copias de este gen. Si el gen viene en dos alelos diferentes, como\(\text{X}^W\) (ojos rojos dominantes, normales) y\(\text{X}^w\) (recesivos, ojos blancos), la mosca hembra puede tener cualquiera de tres genotipos:\(\text{X}^W\text{X}^W\) (ojos rojos),\(\text{X}^W\text{X}^w\) (ojos rojos) y\(\text{X}^w\text{X}^w\) (ojos blancos).
Un macho tiene diferentes posibilidades de genotipo que una hembra. Dado que solo tiene un cromosoma X (emparejado con una Y), solo tendrá una copia de cualquier gen ligado al X. Por ejemplo, en el ejemplo del color del ojo de mosca, los dos genotipos que puede tener un macho son\(\text{X}^W\text{Y}\) (ojos rojos) y\(\text{X}^w\text{Y}\) (ojos blancos). Cualquiera que sea el alelo que herede la mosca macho para un gen ligado al X determinará su apariencia, porque no tiene otra copia genética, incluso si el alelo es recesivo en las hembras. En lugar de homocigotos o heterocigotos, se dice que los machos son hemicigotos para los genes ligados a X.
Podemos ver cómo la vinculación sexual afecta los patrones de herencia al considerar un cruce entre dos moscas, una hembra de ojos blancos (\(\text{X}^w\text{X}^w\)) y un macho de ojos rojos (\(\text{X}^W\text{Y}\)). Si este gen estuviera en un cromosoma no sexual, o autosoma, esperaríamos que toda la descendencia tuviera ojos rojos, porque el alelo rojo es dominante al alelo blanco. Lo que realmente vemos es lo siguiente:
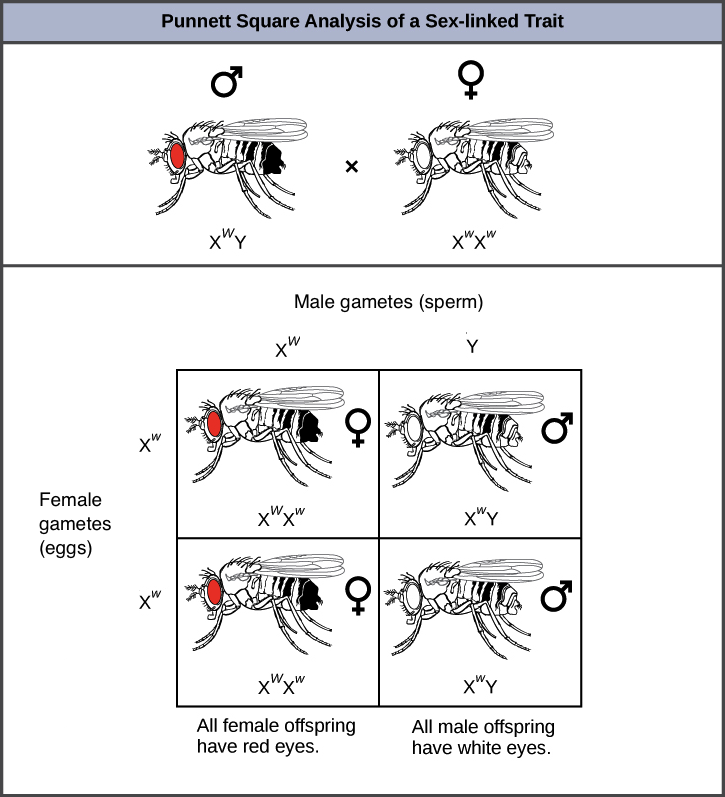
Sin embargo, debido a que el gen está ligado al X, y porque era la madre femenina quien tenía el fenotipo recesivo (ojos blancos), todos los descendientes masculinos —que obtienen su única X de su madre— tienen los ojos blancos (\(\text{X}^w\text{Y}\)). Todas las crías femeninas tienen los ojos rojos porque recibieron dos X, con la\(\text{X}^W\) del padre ocultando la recesiva\(\text{X}^w\) a la madre.
Trastornos genéticos ligados al cromosoma X
Los mismos principios que vemos en el trabajo de las moscas de la fruta se pueden aplicar a la genética humana. En los humanos, los alelos para ciertas afecciones (incluyendo algunas formas de daltonismo, hemofilia y distrofia muscular) están ligados al cromosoma X. Estas enfermedades son mucho más comunes en los hombres que en las mujeres debido a su patrón de herencia ligada al cromosoma X.
¿Por qué es este el caso? Exploremos esto usando un ejemplo en el que una madre es heterocigota para un alelo causante de enfermedad. Se dice que las mujeres heterocigóticas para los alelos de enfermedades son portadoras, y por lo general no muestran ningún síntoma por sí mismas. Los hijos de estas mujeres tienen un 50% de probabilidad de contraer el trastorno, pero las hijas tienen pocas posibilidades de contraer el trastorno (a menos que el padre también lo tenga), y en cambio tendrán un 50% de posibilidades de ser portadoras.
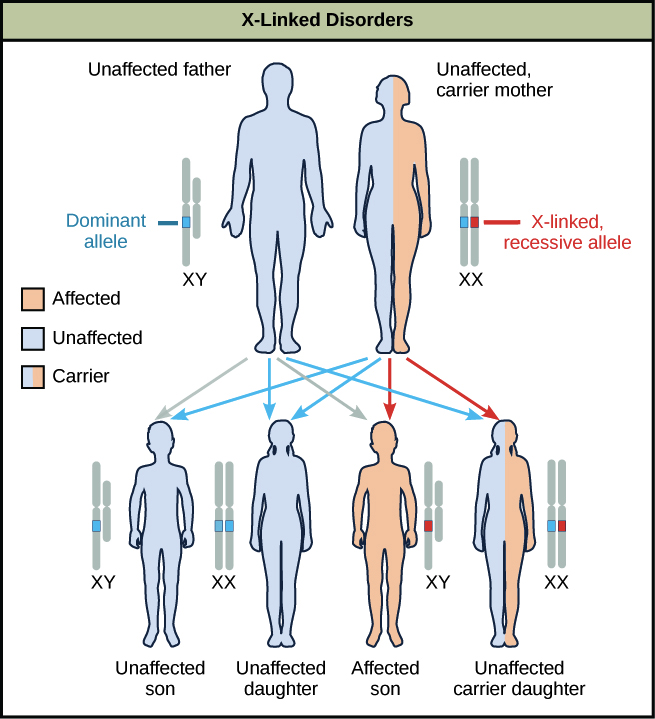
¿Por qué es este el caso? Los rasgos recesivos ligados al X aparecen con más frecuencia en los machos que en las hembras porque, si un macho recibe un alelo “malo” de su madre, no tiene ninguna posibilidad de obtener un alelo “bueno” de su padre (que proporciona una Y) para ocultar el malo. Las hembras, por otro lado, suelen recibir un alelo normal de sus padres, evitando que se exprese el alelo de la enfermedad.
Estudio de caso: Hemofilia
Veamos un ejemplo del cuadrado de Punnett usando un trastorno humano ligado al X: la hemofilia, una afección recesiva en la que la sangre de una persona no se coagula adecuadamente 13. Una persona con hemofilia puede tener sangrado severo, incluso potencialmente mortal, por solo un pequeño corte.
La hemofilia es causada por una mutación en cualquiera de los dos genes, ambos ubicados en el cromosoma X. Ambos genes codifican proteínas que ayudan a coagular la sangre 14. Centrémonos en solo uno de estos genes, llamando al alelo funcional\(\text{X}^H\) y al alelo de la enfermedad\(\text{X}^h\).
En nuestro ejemplo, una mujer heterocigótica para los alelos normales y hemofilia (\(\text{X}^H\text{X}^h\)) tiene hijos con un hombre que es hemicigoto para la forma normal (\(\text{X}^H\text{Y}\)). Ambos padres tienen coagulación sanguínea normal, pero la madre es portadora. ¿Cuál es la probabilidad de que sus hijos e hijas tengan hemofilia?
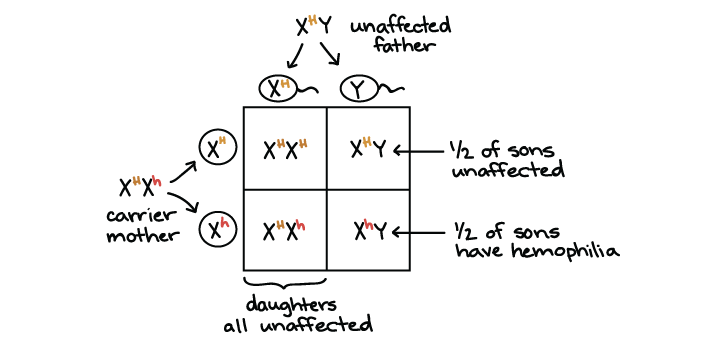
Como la madre es portadora, pasará el alelo de hemofilia (\(\text{X}^h\)) a la mitad de sus hijos, tanto niños como niñas.
- Ninguna de las hijas tendrá hemofilia (probabilidad cero del trastorno). Eso es porque, para tener el trastorno, deben obtener un\(\text{X}^h\) alelo tanto de su madre como de su padre. Hay 0 posibilidades de que las hijas obtengan un\(\text{X}^h\) alelo de su padre, por lo que su probabilidad general de tener hemofilia es cero.
- Los hijos obtienen una Y de su padre en lugar de una X, por lo que su única copia del gen de coagulación de la sangre proviene de su madre. La madre es heterocigota, por lo que la mitad de los hijos, en promedio, obtendrán un\(\text{X}^h\) alelo y tendrán hemofilia (1/2 probabilidad del trastorno).
- [¿Puede una mujer tener hemofilia alguna vez?]
-
Sí, es posible que una mujer tenga una afección recesiva, ligada al cromosoma X como la hemofilia. Sin embargo, debe obtener dos copias recesivas del gen ligado al X (una de cada progenitor) para tener la afección. Las probabilidades de esto son mucho menores que las probabilidades de que un hombre obtenga solo un alelo recesivo de enfermedad de su madre.
Comprueba tu comprensión
Consulta\(\PageIndex{1}\)
- [Pista]
-
Debido a que la hemofilia es un trastorno recesivo, una mujer debe recibir dos alelos de enfermedad (uno en cada cromosoma X) para mostrar la enfermedad. Así, debe obtener un alelo de enfermedad tanto de su madre como de su padre.
Dado que la hemofilia es un trastorno ligado al X, los machos son hemicigotos para el gen relacionado con la hemofilia (tienen solo un alelo y muestran el fenotipo asociado a ese alelo). Para tener un alelo de hemofilia que pueda transmitirse a la descendencia, un macho debe ser él mismo hemofílico. Los pares de padres en los que el macho no es hemofílico no pueden producir una hija hemofílica, salvo eventos raros como mutaciones espontáneas en la línea germinal o durante el desarrollo del embrión.
Las mujeres que son homocigóticas para el alelo de la hemofilia (hemofilia) o heterocigóticas para el alelo de la hemofilia (portadoras no afectadas) pueden transmitir un alelo de hemofilia a sus crías. Los pares de padres en los que la hembra no es ni hemofílica ni portadora no pueden producir una hija hemofílica (nuevamente, salvo raros eventos de mutación espontánea).
De las parejas anteriores, una madre portadora y un padre hemofílico son los que tienen más probabilidades de tener una hija hemofílica.
Colaboradores y Atribuciones
- [Atribución y referencias]
-
Atribución:
Este artículo es un derivado modificado de "Características y rasgos”, por OpenStax College, Biology, CC BY 4.0. Descarga gratis el artículo original en http://cnx.org/contents/185cbf87-c72e-48f5-b51e-f14f21b5eabd@10.53.
Este artículo está bajo una licencia CC BY-NC-SA 4.0.
Obras citadas:
-
Cromosoma X. (2015, 11 de diciembre). Recuperado el 11 de diciembre del 2015 de Wikipedia: https://en.wikipedia.org/wiki/X_chromosome.
-
Reece, J. B., Urry, L. A., Caín, M. L., Wasserman, S. A., Minorsky, P. V., y Jackson, R. B. (2011). La base cromosómica del sexo. En biología Campbell (10a ed., pp. 296-297). San Francisco, CA: Pearson.
-
Cromosoma X. (2012). En Genética hogar de referencia. Recuperado de http://ghr.nlm.nih.gov/chromosome/X.
-
Cromosoma Y. (2010). En Genética hogar de referencia. Recuperado de http://ghr.nlm.nih.gov/chromosome/Y.
-
Meiosis. (2015). En HHMI biointeractivo. Recuperado de http://www.hhmi.org/biointeractive/meiosis.
-
SRY. (2015). En Genética hogar de referencia. Recuperado de http://ghr.nlm.nih.gov/gene/SRY.
-
46, XX trastorno testicular del desarrollo sexual. (2008). En Genética hogar de referencia. Recuperado de https://ghr.nlm.nih.gov/condition/46xx-testicular-disorder-of-sex-development.
-
Síndrome de Swyer. (2015). En Genética hogar de referencia. Recuperado de https://ghr.nlm.nih.gov/condition/swyer-syndrome.
-
King, V., Goodfellow, P. N., Pearks Wilkerson, A. J., Johnson, W. E., O'Brien, S. J., y Pecon-Slattery, J. Evolución del gen determinante masculino SRY dentro de la familia felina Felidae. Genética, 175 (4), 1855-1867. http://dx.doi.org/10.1534/genetics.106.066779.
-
Dimorfismo. (2015). En diccionario Fino. Recuperado de http://www.finedictionary.com/dimorphism.html.
-
Bachtrog, Doris. (2013). Evolución del cromosoma Y: Perspectivas emergentes sobre los procesos de degeneración del cromosoma Y. Nature Opiniones Genética, 14 (2), 113-124. http://dx.doi.org/10.1038/nrg3366. Recuperado de http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4120474/.
-
Gilbert, S. F. (2000). Determinación cromosómica del sexo en Drosophila. En _Biología del Desarrollo (6ª ed.). Sunderland, MA: Asociados Sinauer. Recuperado de www.ncbi.nlm.nih.gov/books/nbk10025/.
-
Hemofilia. (2015). En Genética hogar de referencia. Recuperado de https://ghr.nlm.nih.gov/condition/hemophilia.
-
Kimball, J. W. (2015, 23 de diciembre). Cromosomas sexuales. En las páginas de biología de Kimball. Recuperado el 27 de julio del 2016 de http://www.biology-pages.info/S/SexChromosomes.html.
Referencias adicionales:
Bachtrog, Doris. (2013). Evolución del cromosoma Y: Perspectivas emergentes sobre los procesos de degeneración del cromosoma Y. Nature Opiniones Genética, 14 (2), 113-124. http://dx.doi.org/10.1038/nrg3366. Recuperado de http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4120474/.
Bergmann, D. C. (2011). Notas de conferencias sobre genética. Biosci41, Universidad de Stanford.
Bourke, A. F. G. y Franks, N. R. (1995). Relación y valor reproductivo en los himenópteros sociales. En Evolución social en hormigas (p. 78). Princeton, NJ: Prensa de la Universidad de Princeton.
Bowen, R. A. (2000, 17 de agosto). Preparando un cariotipo. En Genética general y médica. Recuperado de http://www.vivo.colostate.edu/hbooks/genetics/medgen/chromo/cytotech.html.
Carvalho, A. B., Koerich, L. B., y Clark, A. G. (2009). Origen y evolución de los cromosomas Y: cuentos de Drosophila. Tendencias en Genética, 25 (6), 270-277. http://dx.doi.org/10.1016/j.tig.2009.04.002. Recuperado de http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2921885/.
Deeb, S. S. y Motulsky, A. G. (2015, 5 de febrero). Defectos de visión de color rojo-verde. En GenereViews. Recuperado de http://www.ncbi.nlm.nih.gov/books/NBK1301/.
Factores genéticos y hormonas que determinan el género. (2007, 27 de junio). En Embriología Humana: Organogénesis. Recuperado de http://www.embryology.ch/anglais/ugenital/molec02.html.
Haplodiploidía. (2015, 7 de noviembre). Recuperado el 11 de diciembre del 2015 de Wikipedia: https://en.wikipedia.org/wiki/Haplodiploidy.
Kimball, J. W. (2015, 23 de diciembre). Cromosomas sexuales. En las páginas de biología de Kimball. Recuperado el 27 de julio del 2016 de http://www.biology-pages.info/S/SexChromosomes.html.
Krempels, D. M. (n.d.). La genética de los gatos calicó. Recuperado de www.bio.miami.edu/dana/dox/calico.html.
Colegio OpenStax, Biología. (2015, 13 de mayo). Bases cromosómicas de los trastornos hereditarios. En OpenStax CNX. Recuperado de http://cnx.org/contents/185cbf87-c72e-48f5-b51e-f14f21b5eabd@9.85:65/Chromosomal-Basis-of-Inherited.
Colegio OpenStax, Biología. (2015, 13 de mayo). Teoría cromosómica y vinculación genética. En OpenStax CNX. Recuperado a partir de http://cnx.org/contents/185cbf87-c72e-48f5-b51e-f14f21b5eabd@9.85:64/Chromosomal-Theory-and-Genetic
OPN1LW. (2015). En Genética hogar de referencia. Recuperado de http://ghr.nlm.nih.gov/gene/OPN1LW.
Región pseudoautosómica. (2015, 2 de septiembre). Recuperado el 16 de diciembre del 2015 de Wikipedia: https://en.wikipedia.org/wiki/Pseudoautosomal_region.
Purves, W. K., Sadava, D. E., Orians, G. H., y Heller, H.C. (2003). Determinación del sexo y herencia ligada al sexo. En la vida: La ciencia de la biología (7ª ed., pp. 125-144). Sunderland, MA: Asociados Sinauer.
Reece, J. B., Urry, L. A., Caín, M. L., Wasserman, S. A., Minorsky, P. V., y Jackson, R. B. (2011). Los genes ligados al sexo exhiben patrones únicos de herencia. En biología Campbell (10a ed., pp. 205-209). San Francisco, CA: Pearson.
Factor determinante de testículos. (2015, 10 de noviembre). Recuperado el 11 de diciembre del 2015 de Wikipedia: https://en.wikipedia.org/wiki/Testis_determining_factor.
Sistema de determinación de sexo X0 (2015, 20 de julio). Recuperado el 11 de diciembre del 2015 de Wikipedia: https://en.wikipedia.org/wiki/X0_sex-determination_system.
Sistema de determinación del sexo XY. (2015, 19 de noviembre). Recuperado el 11 de diciembre del 2015 de Wikipedia: https://en.wikipedia.org/wiki/XY_sex-determination_system.
Sistema de determinación de sexo ZW. (2015, 16 de noviembre). Recuperado el 11 de diciembre del 2015 de Wikipedia: https://en.wikipedia.org/wiki/ZW_sex-determination_system.
Cigosidad. (2015, 29 de noviembre). Recuperado el 11 de diciembre del 2015 de Wikipedia: https://en.wikipedia.org/wiki/Zygosity.
-