
9: Crecimiento Microbiano
- Page ID
- 56089
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Siempre con las condiciones adecuadas (comida, temperatura correcta, etc) los microbios pueden crecer muy rápidamente. Dependiendo de la situación, esto podría ser algo bueno para los humanos (levadura que crece en mosto para hacer cerveza) o algo malo (bacterias que crecen en la garganta causando faringitis estreptocócica). Es importante tener conocimiento de su crecimiento, para que podamos predecir o controlar su crecimiento en condiciones particulares. Si bien el crecimiento de los organismos muticelulares se mide típicamente en términos del aumento en el tamaño de un solo organismo, el crecimiento microbiano se mide por el aumento de la población, ya sea midiendo el aumento en el número de células o el aumento en la masa general.
División Bacteriana
Las bacterias y las arqueas se reproducen asexualmente solo, mientras que los microbios eucarióticos pueden participar en la reproducción sexual o asexual. Las bacterias y las arqueas se involucran más comúnmente en un proceso conocido como fisión binaria, donde una sola célula se divide en dos células del mismo tamaño. Otros procesos menos comunes pueden incluir fisión múltiple, gemación y producción de esporas.
El proceso comienza con el alargamiento celular, lo que requiere un agrandamiento cuidadoso de la membrana celular y la pared celular, además de un aumento en el volumen celular. La célula comienza a replicar su ADN, en preparación para tener dos copias de su cromosoma, una por cada célula recién formada. La proteína ftsZ es esencial para la formación de un tabique, que inicialmente se manifiesta como un anillo en medio de la célula alargada. Después de segregar los nucleoides a cada extremo de la célula alargada, se completa la formación del tabique, dividiendo la célula alargada en dos células hijas de igual tamaño. Todo el proceso o ciclo celular puede tomar tan solo 20 minutos para un cultivo activo de bacterias E. coli.
Curva de Crecimiento
Dado que las bacterias son fáciles de cultivar en el laboratorio, su crecimiento ha sido ampliamente estudiado. Se ha determinado que en un sistema cerrado o cultivo discontinuo (sin alimentos añadidos, sin residuos eliminados) las bacterias crecerán en un patrón predecible, dando como resultado una curva de crecimiento compuesta por cuatro fases distintas de crecimiento: la fase de retraso, la fase exponencial o logarítmica, la fase estacionaria, y la fase de muerte o declive. Adicionalmente, esta curva de crecimiento puede producir tiempo de generación para un organismo en particular, la cantidad de tiempo que tarda la población en duplicarse.
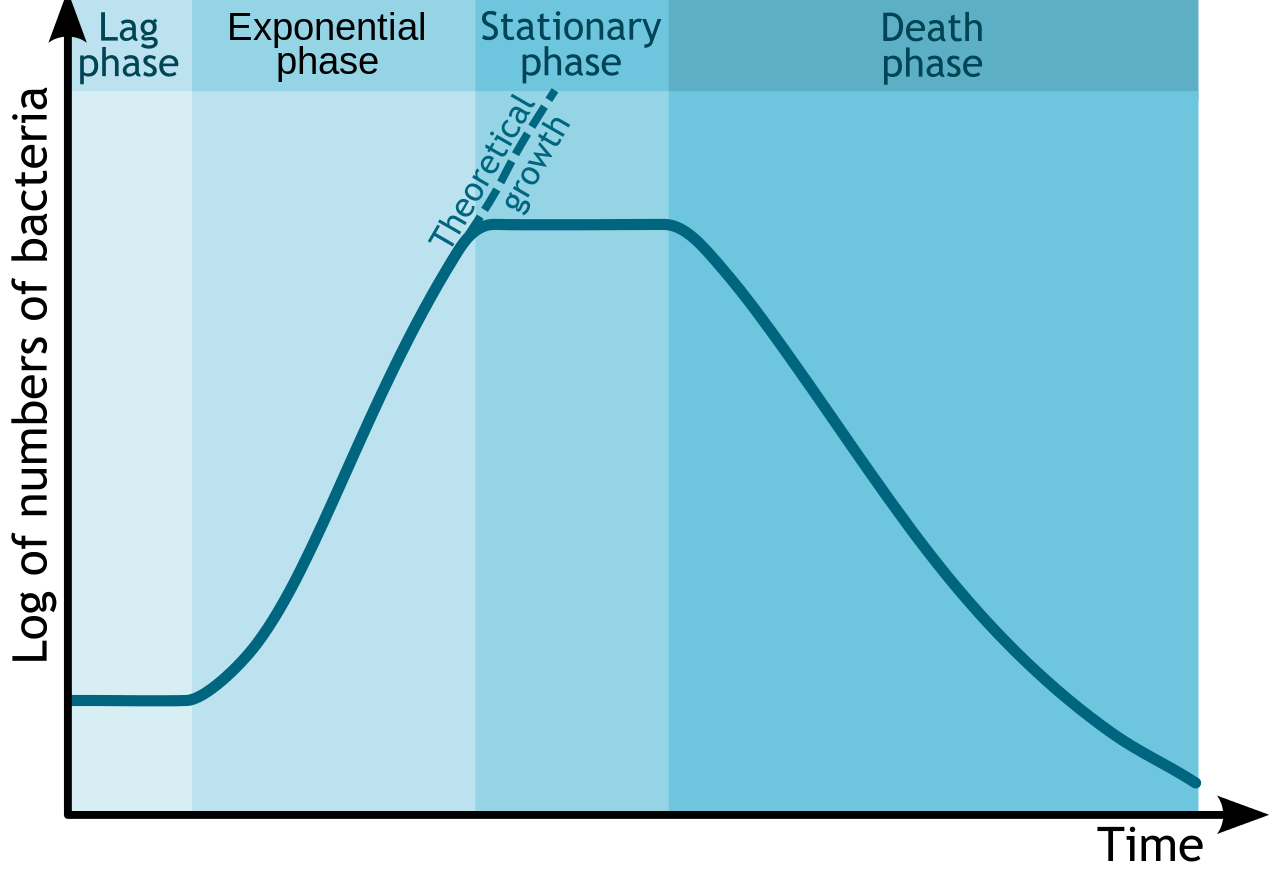
Curva de crecimiento bacteriano. Por Michał Komorniczak. Si utilizas en tu sitio web o en tu publicación mis imágenes (ya sean originales o modificadas), se te solicita que me des datos: Michał Komorniczak (Polonia) o Michal Komorniczak (Polonia). Para mayor información, escribir a mi dirección de correo electrónico: m.komorniczak.pl@gmail.com [CC BY-SA 3.0], vía Wikimedia Commons
Los detalles asociados a cada curva de crecimiento (número de células, longitud de cada fase, rapidez de crecimiento o muerte, cantidad total de tiempo) variarán de organismo a organismo o incluso con diferentes condiciones para un mismo organismo. Pero el patrón de cuatro fases distintas de crecimiento normalmente se mantendrá.
Fase de rezago
La fase de rezago es un periodo de adaptación, donde las bacterias se están ajustando a sus nuevas condiciones. La duración de la fase de retraso puede variar considerablemente, en función de cuán diferentes son las condiciones de las que provienen las bacterias, así como la condición de las propias células bacterianas. Las células que crecen activamente transferidas de un tipo de medio al mismo tipo de medio, con las mismas condiciones ambientales, tendrán el periodo de rezago más corto. Las células dañadas tendrán un largo periodo de rezago, ya que deben repararse por sí mismas antes de que puedan dedicarse a la reproducción.
Por lo general, las células en el periodo de retraso están sintetizando ARN, enzimas y metabolitos esenciales que podrían faltar en su nuevo ambiente (como factores de crecimiento o macromoléculas), además de ajustarse a cambios ambientales como cambios en la temperatura, pH o disponibilidad de oxígeno. También pueden estar realizando cualquier reparación necesaria de células lesionadas.
Fase exponencial o logarítmica
Una vez que las células han acumulado todo lo que necesitan para crecer, proceden a la división celular. La fase exponencial o logarítmica de crecimiento está marcada por duplicaciones predecibles de la población, donde 1 célula se convierte en 2 células, se convierte en 4, se convierte en 8 etc. Las condiciones que son óptimas para las células darán como resultado un crecimiento muy rápido (y una pendiente más pronunciada en la curva de crecimiento), mientras que condiciones inferiores a las ideales dará como resultado un crecimiento más lento. Las células en la fase exponencial de crecimiento son las más sanas y uniformes, lo que explica por qué la mayoría de los experimentos utilizan células de esta fase.
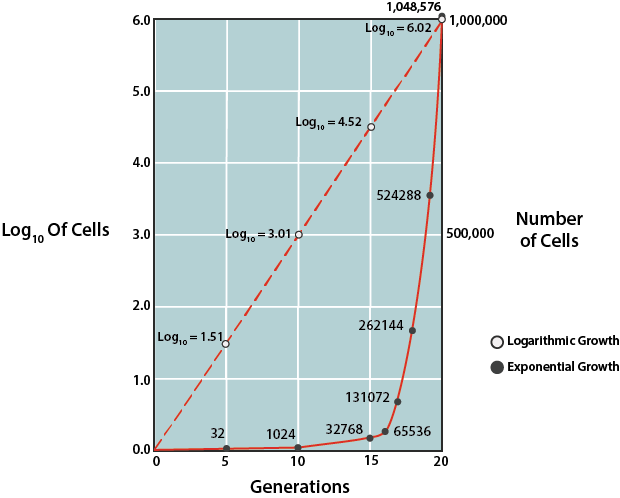
Tasas de Crecimiento Bacteriano.
Debido a la previsibilidad del crecimiento en esta fase, esta fase puede ser utilizada para calcular matemáticamente el tiempo que tarda la población bacteriana en duplicarse en número, conocido como tiempo de generación (g). Esta información es utilizada por microbiólogos en la investigación básica, así como en la industria. Para determinar el tiempo de generación, el logaritmo natural del número de células puede representarse frente al tiempo (donde las unidades pueden variar, dependiendo de la velocidad de crecimiento para la población particular), utilizando una gráfica semilogarítmica para generar una línea con una pendiente predecible.
La pendiente de la línea es igual a 0.301/ g. Alternativamente, se puede confiar en la relación fija entre el número inicial de celdas al inicio de la fase exponencial y el número de celdas después de algún período de tiempo, que puede expresarse mediante:
\[N = N_02^{n}\]
donde\(N\) es la concentración celular final,\(N_0\) es la concentración celular inicial, y\(n\) es el número de generaciones que ocurrieron entre el periodo de tiempo especificado.
El tiempo de generación (g) puede ser representado por t/n, siendo t el periodo de tiempo especificado en minutos, horas, días o meses. Así, si se conoce la concentración celular al inicio de la fase exponencial de crecimiento y la concentración celular después de algún periodo de tiempo de crecimiento exponencial, se puede calcular el número de generaciones. Entonces, utilizando la cantidad de tiempo que se permitió que el crecimiento continuara (t), se puede calcular g.
Fase Estacionaria
Todas las cosas buenas deben llegar a su fin (¡de lo contrario las bacterias equivaldrían a la masa de la Tierra en 7 días!). En algún momento la población bacteriana se queda sin un nutriente/químico esencial o su crecimiento es inhibido por sus propios productos de desecho (es un contenedor cerrado, ¿recuerdas?) o falta de espacio físico, haciendo que las células entren en la fase estacionaria. En este punto, el número de nuevas células que se producen es igual al número de células que mueren o el crecimiento ha cesado por completo, dando como resultado un aplanamiento del crecimiento en la curva de crecimiento.
Fisiológicamente las células se vuelven bastante diferentes en esta etapa, ya que intentan adaptarse a sus nuevas condiciones de inanición. Las pocas células nuevas que se producen son de menor tamaño, con bacilos volviéndose casi esféricos en forma. Su membrana plasmática se vuelve menos fluida y permeable, con moléculas más hidrofóbicas en la superficie que promueven la adhesión y agregación celular. El nucleoide se condensa y el ADN se une con proteínas de unión al ADN de células hambrientas (DPS), para proteger el ADN del daño. Los cambios están diseñados para permitir que la célula sobreviva por un periodo de tiempo más largo en condiciones adversas, mientras se espera que se produzcan condiciones más óptimas (como una infusión de nutrientes). Estas mismas estrategias son utilizadas por las células en ambientes oligotróficos o bajos en nutrientes. Se ha planteado la hipótesis de que las células en el mundo natural (es decir, fuera del laboratorio) suelen existir por largos periodos de tiempo en ambientes oligotróficos, con solo infusiones esporádicas de nutrientes que las devuelven a un crecimiento exponencial por periodos de tiempo muy breves.
Durante la fase estacionaria las células también son propensas a producir metabolitos secundarios, o metabolitos producidos después del crecimiento activo, como los antibióticos. Las células que son capaces de producir una endospora activarán los genes necesarios durante esta etapa, con el fin de iniciar el proceso de esporulación.
Fase de muerte o declive
En la última fase de la curva de crecimiento, la fase de muerte o declive, el número de células viables disminuye de manera predecible (o exponencial). La pendiente de la pendiente corresponde a la rapidez con la que las células están perdiendo viabilidad. Se piensa que las condiciones de cultivo se han deteriorado hasta un punto en el que las células se ven irreparablemente dañadas, ya que las células recolectadas de esta fase no logran mostrar crecimiento cuando se transfieren a medio fresco. Es importante señalar que si se está midiendo la turbidez de un cultivo como una forma de determinar la densidad celular, las mediciones podrían no disminuir durante esta fase, ya que las células aún podrían estar intactas.
Se ha sugerido que las células que se cree que están muertas podrían ser revividas en condiciones específicas, una condición descrita como viable pero no cultivable (VBNC). Este estado podría ser de importancia para los patógenos, donde entran en un estado de metabolismo muy bajo y falta de división celular, sólo para reanudar el crecimiento en un momento posterior, cuando las condiciones mejoran.
También se ha demostrado que el 100% de muerte celular es poco probable, para cualquier población celular, ya que las células mutan para adaptarse a sus condiciones ambientales, por muy duras que sean. A menudo se observa un efecto de cola, donde una pequeña población de las células no puede ser eliminada. Además, estas células podrían beneficiarse de la muerte de sus compañeras, las cuales aportan nutrientes al ambiente a medida que lisan y liberan su contenido celular.
Palabras clave
fisión binaria, fisión múltiple, gemación, esporas, ciclo celular, sistema cerrado, cultivo discontinuo, curva de crecimiento, fase lag, fase exponencial o logarítmica, tiempo de generación (g), N, N0, n, t, fase estacionaria, proteínas de unión a ADN de células privadas de hambre (DPS), oligotróficas, secundarias metabolitos, fase de muerte o declive, viables pero no cultivables (VBNC).
Preguntas/Objetivos Esenciales
- ¿Cómo se mide el crecimiento en poblaciones microbianas?
- ¿En qué se diferencian los eucariotas y las bacterias/arqueas en sus métodos reproductivos?
- ¿Cuáles son los pasos de la fisión binaria? ¿Qué sucede en cada paso?
- Conoce cómo es la curva de crecimiento de un organismo cultivado en un sistema cerrado. Conocer las diversas etapas y lo que está ocurriendo en cada etapa, fisiológicamente. ¿Qué puede influir en la fase de retraso? ¿Cuáles son las 2 explicaciones diferentes para la pérdida celular en la fase de muerte o senescencia?
- Entender el tiempo de generación y cómo se puede determinar en un gráfico de número logarítmico de celdas vs. tiempo. Conozca la ventaja de trazar el número logarítmico de celdas frente al tiempo en lugar del número de celdas frente al tiempo. ¿Qué factores afectan el tiempo de generación de un organismo?
- Problema de práctica: Seis Staphylococcus aureus son inoculados en un pastel de crema de manos de un chef pastelero. El tiempo de generación de S. aureus en pastel de crema a temperatura ambiente es de 30 minutos. a) ¿Cuántos S. aureus hay en el pastel después de 4 horas a TA? b) ¿Pasadas las 24 horas?
Preguntas Exploratorias (OPCIONAL)
- ¿En qué situación beneficiaría la ocurrencia de VBNC a las células? ¿Cómo podría esto representar una amenaza para la salud pública?