
13: Quimioorganotrofia
- Page ID
- 56092
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)La quimioorganotrofia es un término utilizado para denotar la oxidación de productos químicos orgánicos para producir energía. En otras palabras, una sustancia química orgánica sirve como donador de electrones inicial. El proceso se puede realizar en presencia o ausencia de oxígeno, dependiendo de lo que esté disponible para una célula y de si tienen o no las enzimas para tratar los subproductos tóxicos del oxígeno.
Respiración aeróbica
Para comenzar, centrémonos en el catabolismo de los compuestos orgánicos cuando ocurre en presencia de oxígeno. En otras palabras, el oxígeno se está utilizando como aceptor final de electrones. Cuando el proceso utiliza glucólisis y el ciclo del ácido tricarboxílico (TCA) para oxidar completamente un compuesto orgánico hasta CO 2, se conoce como respiración aeróbica. Esto genera la mayor cantidad de ATP para una célula, dada la gran cantidad de distancia entre el donante de electrones inicial (glucosa) y el aceptor de electrones final (oxígeno), así como la gran cantidad de electrones que tiene que donar la glucosa.
Fuentes de Energía Orgánicas
En la quimioorganotrofia, la energía se deriva de la oxidación de un compuesto orgánico. Hay muchos compuestos orgánicos diferentes disponibles para una célula, como proteínas, polisacáridos y lípidos. Pero las vías celulares están dispuestas de tal manera que aumentan la eficiencia metabólica. Así, la célula emana las reacciones en algunas vías comunes. Por convención, la glucosa se utiliza como molécula de partida para describir cada proceso.
Glicólisis
La glucólisis es una vía casi universal para el catabolismo de la glucosa al piruvato. La vía se divide en dos partes: la parte I, que se centra en las modificaciones a la glucosa de azúcar de 6 carbonos, y la parte II, donde el compuesto de 6 carbonos se divide en dos moléculas de 3 carbonos, produciendo una vía bifurcada. La Parte I en realidad requiere energía en forma de 2 moléculas de ATP, para fosforilar o activar el azúcar. La Parte II es la fase conservadora de energía de la reacción, donde se generan 4 moléculas de ATP por fosforilación a nivel de sustrato, donde una molécula de alta energía transfiere directamente un P i a ADP.
El rendimiento neto de energía de la glucólisis es de 2 moléculas de ATP por cada molécula de glucosa. Además, se reducen 2 moléculas del portador NAD+, formando NADH. En la respiración aeróbica, estos electrones serán finalmente transferidos por el NADH a una cadena de transporte de electrones, permitiendo que la célula capture más energía. Por último, se producen 2 moléculas del compuesto de 3 carbonos piruvato, que pueden oxidarse aún más para capturar más energía para la célula.
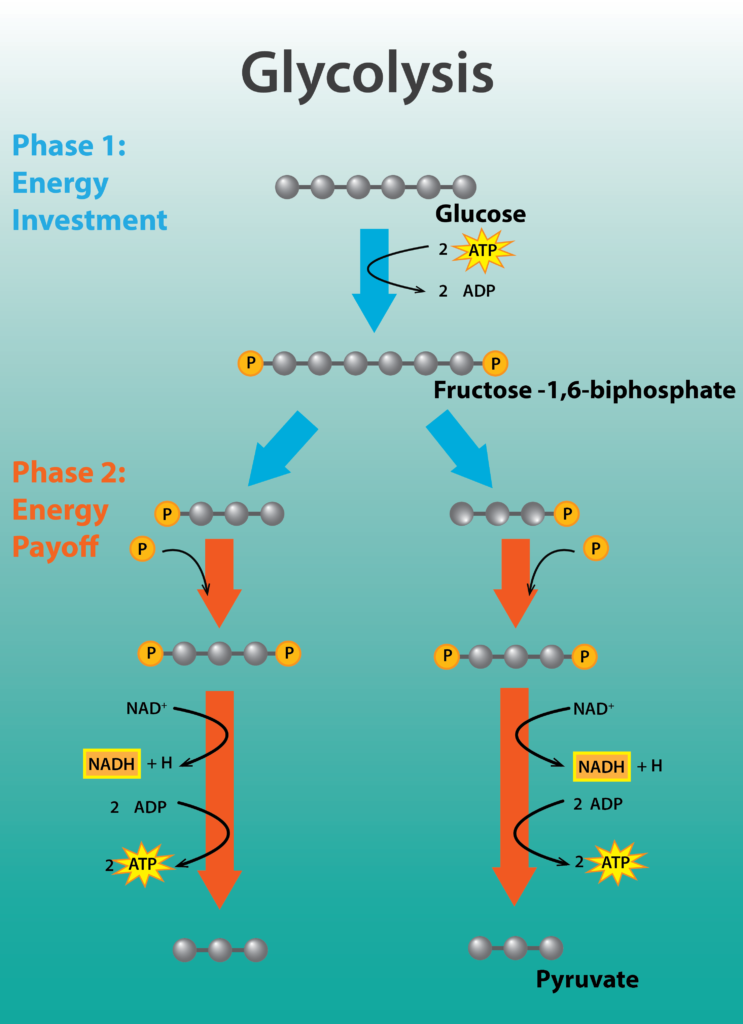
Glicólisis.
Ciclo del ácido tricarboxílico (TCA)
El ciclo del ácido tricarboxílico (TCA) se recoge al final de la glucólisis, con el fin de oxidar completamente cada molécula de piruvato hasta 3 moléculas de CO 2, como ocurre en la respiración aeróbica. Comienza con un tipo de reacción de conexión antes de que las moléculas puedan entrar en el ciclo propiamente dicho. La reacción de conexión reduce 1 molécula de NAD+ a NADH por cada molécula de piruvato, en el proceso de elaboración de citrato.
El citrato ingresa a la parte del ciclo real del proceso, experimentando una serie de oxidaciones que producen muchos productos diferentes, muchos de ellos importantes metabolitos precursores para otras vías. A medida que se liberan electrones, los portadores se reducen, produciendo 3 moléculas de NADH y 1 molécula de FADH 2 por cada molécula de piruvato. Además, se genera 1 molécula de GTP (que puede considerarse como una molécula equivalente a ATP) por fosforilación a nivel de sustrato.
Teniendo en cuenta que hubo dos moléculas de piruvato generadas a partir de la glucólisis, el rendimiento neto del ciclo de TCA y su reacción de conexión son: 2 moléculas de GTP, 8 moléculas de NADH y 2 moléculas de FADH 2. Pero, ¿de dónde viene la ATP? Hasta ahora solo tenemos el rendimiento neto de 2 moléculas de glucólisis y las 2 moléculas de equivalentes de ATP (es decir, GTP) del ciclo de TCA. Aquí es donde entra en juego la cadena de transporte de electrones.
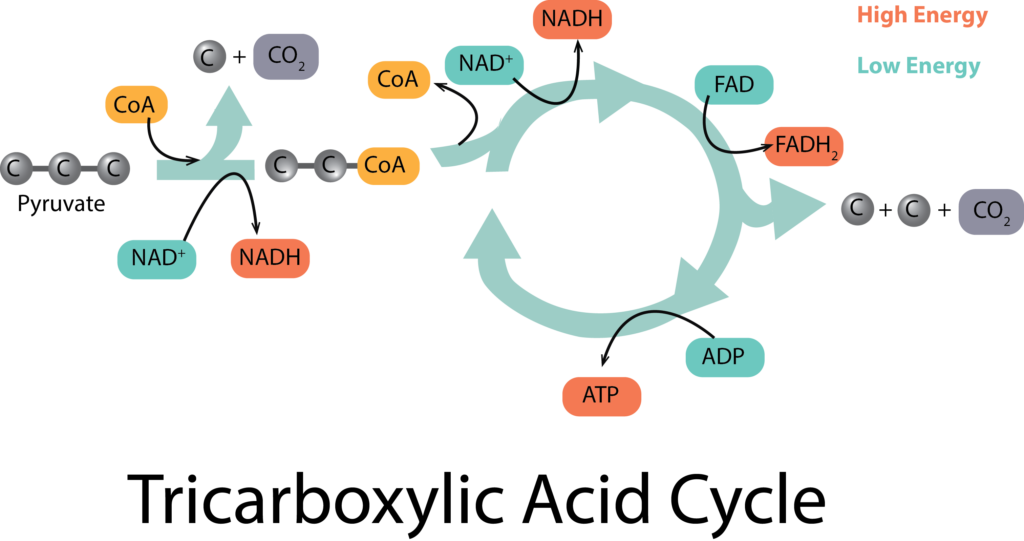
TCA al Fin de la Glicólisis.
Fosforilación oxidativa
La síntesis de ATP a partir del transporte de electrones generado por la oxidación de una fuente de energía química es conocida como fosforilación oxidativa. Ya hemos establecido que los electrones pasan de portador a portador, en orden de su potencial de reducción estándar. También hemos establecido que algunos portadores aceptan electrones y protones, mientras que otros aceptan electrones únicamente. ¿Qué pasa con los protones no aceptados? ¿Y cómo genera esto ATP para la célula? ¡Bienvenido al maravilloso mundo de la fuerza motriz de protones (PMF) y la ATP sintasa!
Fuerza Motiva de Protones
Los protones que no son aceptados por los portadores de electrones migran hacia afuera, para revestir la parte externa de la membrana. Para bacterias y arqueas, esto significa revestir la membrana celular y explica la importancia para la carga negativa de la célula.
A medida que se acumulan los protones cargados positivamente, se desarrolla un gradiente de concentración de protones. Esto da como resultado que el citoplasma de la célula sea más alcalino y más negativo, lo que lleva a una diferencia de potencial tanto químico como eléctrico. Esta fuerza motriz protónica (PMF) puede ser utilizada para hacer trabajo para la célula, como en la rotación del flagelo bacteriano o la captación de nutrientes.
ATP sintasa
El PMF también se puede utilizar para sintetizar ATP, con la ayuda de una enzima conocida como ATP sintasa (o ATPasa). Esta gran enzima tiene dos componentes, uno que abarca la membrana y otro que se adhiere al citoplasma y sintetiza el ATP. Los protones son impulsados a través del componente que abarca la membrana, generando un par que impulsa la rotación de la porción citoplásmica. Cuando el componente citoplásmico vuelve a su configuración original se une a P i a ADP, generando una molécula de ATP.
Resumen de respiración aeróbica
Después de todo eso, ¿con qué terminó la célula, de usar la respiración aeróbica? Mediante la fosforilación a nivel de sustrato, la célula generó 2 moléculas netas de ATP durante la glucólisis, además de 2 moléculas de equivalentes de ATP del ciclo de TCA. Para portadores reducidos, hubo 2 moléculas de NADH generadas durante la glucólisis, además de 8 moléculas del ciclo de TCA o su reacción de conexión. También hubo 2 moléculas de FADH 2 del ciclo de TCA. Todos esos electrones se pasaron a la ETC (y eventualmente al oxígeno), con el fin de desarrollar un PMF, de manera que la ATP sintasa pudiera generar ATP. ¿Cuánto ATP se genera?
Las investigaciones indican que el proceso no es completamente eficiente y hay alguna “fuga” que ocurre. Las estimaciones actuales son que se generan 2.5 ATP por cada molécula de NADH, mientras que 1.5 ATP se generan por cada molécula de FADH 2. El uso de estos valores permitiría a la célula sintetizar 25 moléculas de ATP de todo el NAD+ que se redujo en el proceso, además de 3 moléculas de ATP del FAD+ que se redujo. Esto llevaría el gran total de ATP máximo producido a 32 (contando el GTP en esa cifra).
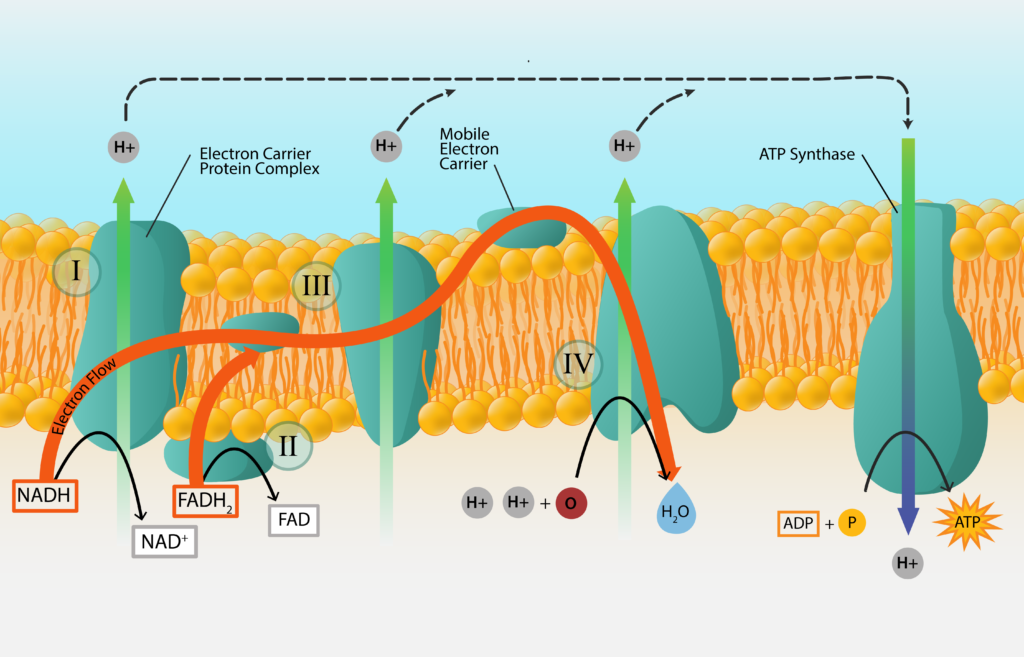
Generación ATP.
Quimioorganotrofia anaerobia
Ciertamente, el oxígeno es un maravilloso aceptor final de electrones, particularmente cuando se combina con la glucosa como donante de electrones inicial. Es parte del par redox más bajo en una torre de electrones, con un potencial electrónico estándar extremadamente positivo. Pero, ¿qué hace un microbio, si el oxígeno no está disponible o carece de las protecciones necesarias frente a los subproductos tóxicos del oxígeno? Centrémonos en la generación de energía en ausencia de oxígeno, utilizando un aceptor de electrones diferente, cuando todavía se está utilizando una sustancia química orgánica como donador de electrones inicial. Los ejemplos de quimioorganotrofia anaerobia incluyen respiración anaeróbica y fermentación.
Respiración anaeróbica
La respiración anaeróbica también comienza con glucólisis y el piruvato puede desviarse al ciclo de TCA, al igual que en la respiración aeróbica. De hecho, la fosforilación oxidativa se utiliza para generar la mayor parte del ATP, lo que significa el uso de una ETC y ATP sintasa. La diferencia clave es que el aceptor de electrones final no será oxígeno.
Hay una variedad de posibles aceptores finales de electrones que pueden ser utilizados en la respiración anaeróbica, permitiendo que los microbios vivan en una amplia variedad de ubicaciones. El mejor aceptor de electrones será el que esté más bajo en la torre de electrones, en forma oxidada (es decir, en el lado izquierdo del par redox). Algunos aceptores de electrones comunes incluyen nitrato (NO3-), hierro férrico (Fe3+), sulfato (SO42-), carbonato (CO32-) o incluso ciertos compuestos orgánicos, como fumarato.
¿Cuánto ATP es generado por la respiración anaeróbica? Eso dependerá del aceptor de electrones final que se utilice. No será tanto como se genera durante la respiración aeróbica, ya que sabemos que el oxígeno está en el mejor aceptor de electrones posible. La selección de un aceptor de electrones distinto del oxígeno empuja a un organismo hacia arriba por la torre de electrones, acortando la distancia entre el donador de electrones y el aceptor, reduciendo la cantidad de ATP producido.
Fermentación
No importa lo que te puedan enseñar en una clase bioquímica, la fermentación y la respiración anaeróbica no son lo mismo, al menos no a un microbiólogo.
La fermentación es catabolismo de la glucosa en ausencia de oxígeno también y tiene algunas similitudes con la respiración anaeróbica. Lo más obvio es que no utiliza oxígeno como aceptor final de electrones. En realidad utiliza piruvato, un compuesto orgánico. La fermentación comienza con la glucólisis, un proceso que ya hemos cubierto, que también inicia tanto la respiración aeróbica como la respiración anaeróbica. ¿Qué rinde? Dos moléculas netas de ATP por fosforilación a nivel de sustrato y 2 moléculas de NADH. Los organismos que hacen respiración aeróbica o anaeróbica utilizarían la fosforilación oxidativa para aumentar su rendimiento de ATP. Los fermentadores, sin embargo, carecen de un ETC o reprimen la síntesis de su ETC cuando no hay oxígeno disponible, por lo que no utilizan el ciclo de TCA en absoluto.
Sin el uso de una ETC (o una PMF o ATP sintasa), no se genera más ATP más allá de lo que se sintetizó durante la glucólisis. Pero los organismos que utilizan la fermentación no pueden simplemente detenerse con la glucólisis, ya que eventualmente todas sus moléculas de NAD+ se reducirían. Para reoxidar este portador de electrones utilizan piruvato como aceptor final de electrones, produciendo una variedad de productos de fermentación como etanol, CO 2 y diversos ácidos.
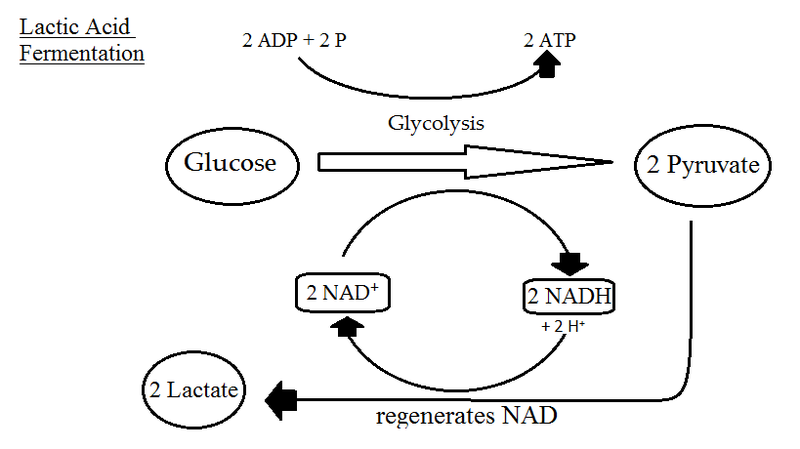
Fermentación de Lactato. Por Sjantoni (Obra propia) [CC BY-SA 3.0], vía Wikimedia Commons
Los productos de fermentación, aunque considerados productos de desecho para la célula, son de vital importancia para los humanos. Nos apoyamos en el proceso de fermentación para producir una variedad de alimentos fermentados (cerveza, vino, pan, queso, tofu), además de utilizar los productos para una variedad de procesos industriales.
Palabras clave
quimioorganotrofia, respiración aeróbica, glucólisis, fosforilación a nivel de sustrato, ciclo de ácido tricarboxílico (TCA), GTP, fosforilación oxidativa, fuerza motriz protónica (PMF), ATP sintasa/ATPasa, respiración anaeróbica, fermentación.
Preguntas de Estudio
- ¿Qué es la quimioorganotrofia?
- En la glucólisis, ¿cuál es el compuesto de partida? ¿Cuántas moléculas de ATP (total y neta) se producen? ¿Cómo se reducen las moléculas de NADH?
- ¿Qué es la fosforilación a nivel de sustrato?
- ¿Cómo reoxidan los organismos NADH, después de la descomposición de la glucosa a piruvato? ¿Por qué es importante para ellos reoxidar el NADH?
- Durante el ciclo de TCA y la reacción de conexión, ¿en qué se descompone la glucosa? ¿Cuántas moléculas de equivalentes de ATP/ATP se forman por la fosforilación del sustrato? ¿Cuántas moléculas de NAD y cuántas moléculas de FAD se reducen?
- ¿Qué obtiene la célula del ciclo TCA, en términos de energía e intermedios?
- En la respiración aeróbica, ¿cómo se reoxida el NADH? ¿Cuál es el ATP máximo por NADH o FADH formado durante esta reoxidación? ¿Cuál es el aceptor de electrones final?
- ¿Qué componentes están involucrados en el transporte de electrones? ¿Qué es una fuerza motriz protónica y qué papel juega en la generación de energía?
- ¿Qué es la fosforilación oxidativa? ¿Dónde específicamente se desprende energía en el transporte de electrones y cómo se conserva esa energía?
- ¿Cómo funciona la ATP sintasa para cosechar la energía conservada?
- ¿Cuántos ATP se forman cuando la glucosa se descompone completamente en la respiración aeróbica bacteriana y de dónde provienen? ¿Qué otros productos se forman?
- ¿Cómo es similar y diferente la respiración anaeróbica de la respiración aeróbica? ¿Cómo se compara el rendimiento energético? ¿Por qué?
- ¿Cómo es la fermentación similar y diferente a la respiración aeróbica y anaeróbica? ¿Cómo se compara el rendimiento energético? ¿Por qué? ¿Cuáles son los productos finales de la fermentación?
- Para cada tipo de metabolismo en este capítulo, ¿cuál es el donante de electrones inicial? ¿Cuál es el aceptor de electrones final? ¿Qué procesos se utilizan para generar energía? ¿Cuál es el rendimiento energético?
Fermentación de Lactato. Por Sjantoni (Obra propia) [CC BY-SA 3.0], vía Wikimedia Commons
Los productos de fermentación, aunque considerados productos de desecho para la célula, son de vital importancia para los humanos. Nos apoyamos en el proceso de fermentación para producir una variedad de alimentos fermentados (cerveza, vino, pan, queso, tofu), además de utilizar los productos para una variedad de procesos industriales.
Palabras clave
quimioorganotrofia, respiración aeróbica, glucólisis, fosforilación a nivel de sustrato, ciclo de ácido tricarboxílico (TCA), GTP, fosforilación oxidativa, fuerza motriz protónica (PMF), ATP sintasa/ATPasa, respiración anaeróbica, fermentación.
Preguntas de Estudio
- ¿Qué es la quimioorganotrofia?
- En la glucólisis, ¿cuál es el compuesto de partida? ¿Cuántas moléculas de ATP (total y neta) se producen? ¿Cómo se reducen las moléculas de NADH?
- ¿Qué es la fosforilación a nivel de sustrato?
- ¿Cómo reoxidan los organismos NADH, después de la descomposición de la glucosa a piruvato? ¿Por qué es importante para ellos reoxidar el NADH?
- Durante el ciclo de TCA y la reacción de conexión, ¿en qué se descompone la glucosa? ¿Cuántas moléculas de equivalentes de ATP/ATP se forman por la fosforilación del sustrato? ¿Cuántas moléculas de NAD y cuántas moléculas de FAD se reducen?
- ¿Qué obtiene la célula del ciclo TCA, en términos de energía e intermedios?
- En la respiración aeróbica, ¿cómo se reoxida el NADH? ¿Cuál es el ATP máximo por NADH o FADH formado durante esta reoxidación? ¿Cuál es el aceptor de electrones final?
- ¿Qué componentes están involucrados en el transporte de electrones? ¿Qué es una fuerza motriz protónica y qué papel juega en la generación de energía?
- ¿Qué es la fosforilación oxidativa? ¿Dónde específicamente se desprende energía en el transporte de electrones y cómo se conserva esa energía?
- ¿Cómo funciona la ATP sintasa para cosechar la energía conservada?
- ¿Cuántos ATP se forman cuando la glucosa se descompone completamente en la respiración aeróbica bacteriana y de dónde provienen? ¿Qué otros productos se forman?
- ¿Cómo es similar y diferente la respiración anaeróbica de la respiración aeróbica? ¿Cómo se compara el rendimiento energético? ¿Por qué?
- ¿Cómo es la fermentación similar y diferente a la respiración aeróbica y anaeróbica? ¿Cómo se compara el rendimiento energético? ¿Por qué? ¿Cuáles son los productos finales de la fermentación?
- Para cada tipo de metabolismo en este capítulo, ¿cuál es el donante de electrones inicial? ¿Cuál es el aceptor de electrones final? ¿Qué procesos se utilizan para generar energía? ¿Cuál es el rendimiento energético?