
15: Fototrofia
- Page ID
- 56049
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Por supuesto, obtener energía de la luz solar, eso es bastante genial. ¿O hace calor? Dejando a un lado las bromas malas, ojalá recuerdes el proceso básico de fotosíntesis de clases anteriores, donde podrían haber hablado de lo que sucede en las plantas. ¿El camino Z? ¿Algo? No se asuste — ya hablaremos de ello. Junto con los otros tipos de fototrofia que utilizan los microbios.
Fotoautótrofos vs fotoheterótrofos
La fototrofia (o “comer luz”) se refiere al proceso por el cual la energía del sol es capturada y convertida en energía química, en forma de ATP. El término fotosíntesis se usa con mayor precisión para describir organismos que convierten la luz solar en ATP (la “reacción de la luz”) pero luego también proceden a usar el ATP para fijar el dióxido de carbono en compuestos orgánicos (el ciclo Calvino). Estos organismos son los fotoautótrofos. En el mundo microbiano, también hay fotoheterótrofos, organismos que convierten la luz solar en ATP pero utilizan compuestos orgánicos prefabricados disponibles en el medio ambiente. El ATP podría entonces ser utilizado para otros fines.
Pigmentos
Para convertir la energía de la luz solar en ATP, los organismos utilizan pigmentos sensibles a la luz. Las plantas y las algas utilizan clorofilas, que también son utilizadas por las cianobacterias. Las clorofilas son de color verde, debido a que absorben longitudes de onda rojas y azules (≈675 nm y 430 nm) y transmiten luz verde. Las bacterias púrpura y verde tienen bacterioclorofilas, que absorben longitudes de onda más altas (≈870 nm) que las clorofilas, permitiendo que diferentes fototrofos ocupen el mismo ambiente sin competir entre sí.
Los fototrofos también pueden contener pigmentos accesorios, como los carotenoides y las ficobiliproteínas. Los carotenoides, que absorben la luz azul (400-550 nm), suelen ser de color amarillo, naranja o rojo. Las ficobiliproteínas pueden dividirse en dos grupos: ficoeritrina, que transmite un color rojo, y ficocianina, que transmite un color azul. Los pigmentos accesorios pueden servir para ampliar el rango de longitud de onda de la luz que se absorbe, permitiendo una mejor utilización de la luz disponible. Además, estos pigmentos pueden servir una función protectora para el organismo al actuar como antioxidante.
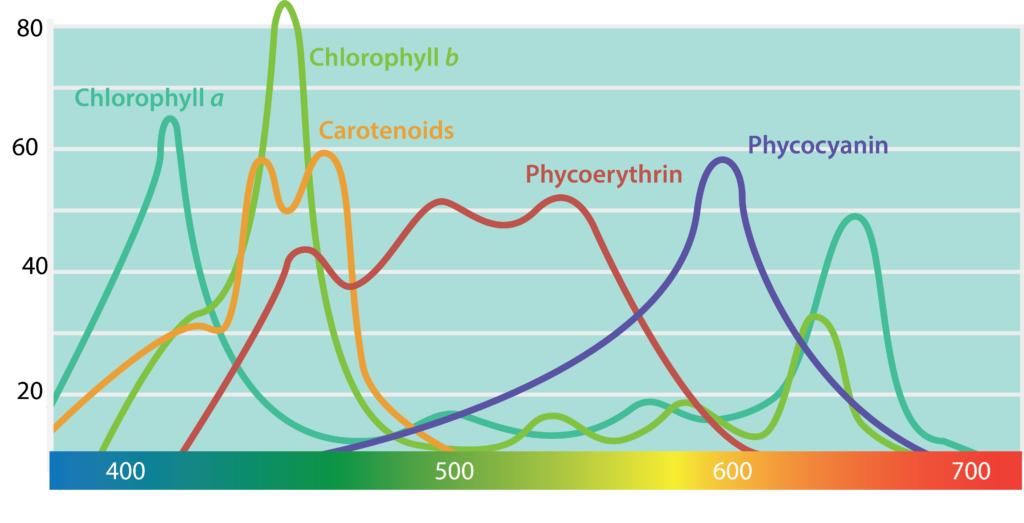
Pigmento Fototrófico.
En bacterias y arqueas, los pigmentos fototróficos se alojan dentro de invaginaciones de la membrana celular o dentro de un clorosoma. Los pigmentos de recolección de luz forman antenas, que canalizan la luz a otras moléculas en los centros de reacción, que en realidad realizan la conversión de la energía lumínica en ATP.
Fotofosforilación en general
Para cualquier organismo, el proceso general de fototrofia va a ser el mismo. Una antena de fotosistema absorbe la luz y canaliza la energía a un centro de reacción, específicamente a un par especial de moléculas de clorofila/bacterioclorofila. Las moléculas se excitan, cambiando a un potencial de reducción más negativo (es decir, saltando la torre de electrones). Los electrones pueden entonces pasar a través de una cadena de transporte de electrones de portadores, como la ferredoxina y los citocromos, permitiendo el desarrollo de una fuerza motriz protónica. Los protones son llevados de vuelta a través de la membrana plasmática a través de ATPasa, generando ATP en el proceso. Dado que la energía original del proceso vino de la luz solar, a diferencia de un químico, el proceso se llama fotofosforilación. Si los electrones son devueltos al par especial de moléculas de clorofila/bacterioclorofila (fotofosforilación cíclica), el proceso puede repetirse una y otra vez. Si los electrones se desvían a otra parte, como para la reducción de NAD (P) (fotofosforilación no cíclica), entonces se debe utilizar una fuente de electrones externa para reponer el sistema.
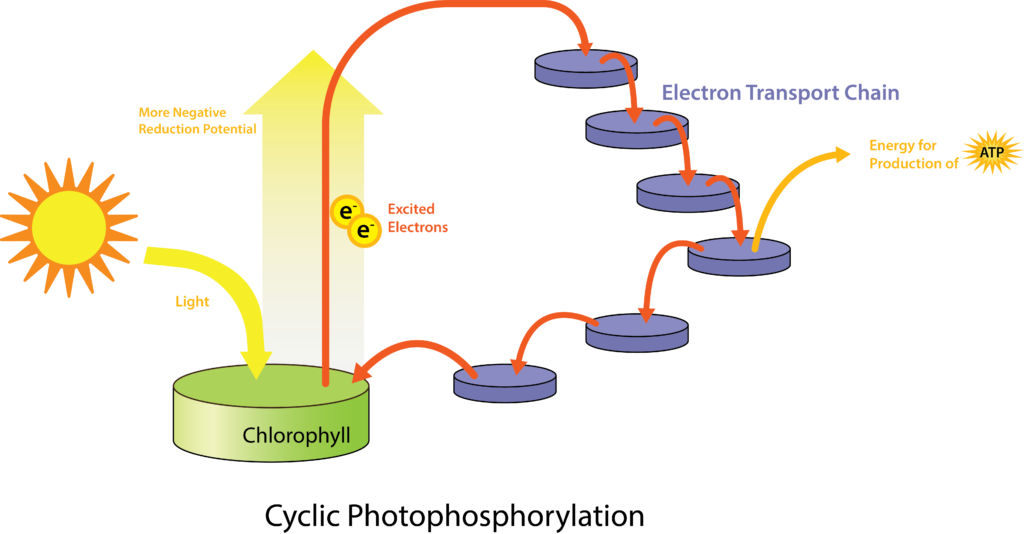
Fototrofia anoxígena
Bacteria fototrófica púrpura
Las bacterias fototróficas moradas se involucran en la fototrofia anoxígena, lo que indica que no generan oxígeno durante el proceso. Tienen un solo fotosistema con bacterioclorofila, lo que les permite utilizar fotofosforilación cíclica como se describió anteriormente para la formación de ATP. Pero si una bacteria púrpura quiere crecer como fotoautótrofa, también necesitará poder reductor en forma de NAD (P) H.
El centro de reacción de la bacteria púrpura (conocida como P870) tiene un E0' de +0.5V. Después de ser alcanzado por un fotón de luz, el potencial cambia a -1.0V, lo que es insuficiente para reducir NAD (P) con su E0' de -0.32V. Por lo tanto, las bacterias púrpuras autótrofas deben participar en un proceso conocido como flujo inverso de electrones, utilizando energía de la fuerza motriz protónica para impulsar electrones por la torre de electrones. Adicionalmente, deben encontrar un donador de electrones externo para reponer los electrones ahora desviados a NAD (P). Típicamente los electrones provienen de H2S o azufre elemental, con diversos subproductos de azufre producidos.
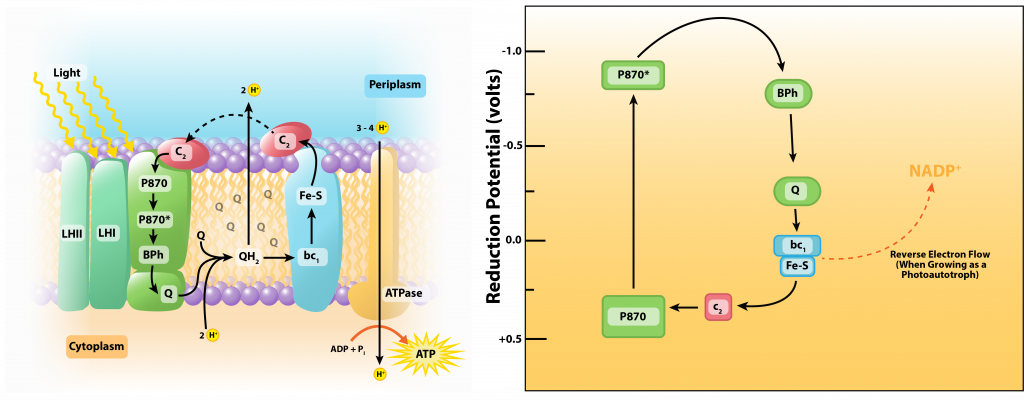
Fotofosforilación en Bacterias Moradas.
En presencia de compuestos orgánicos, las bacterias púrpuras suelen existir como fotoheterótrofos, utilizando la fotofosforilación cíclica para generar ATP y obtener sus compuestos orgánicos del ambiente. Esto elimina la necesidad de utilizar flujo inverso de electrones, un proceso energéticamente desfavorable, así como la necesidad de donantes de electrones externos.
Bacterias fototróficas verdes
Las bacterias fototróficas verdes también participan en la fototrofia anoxigénica, utilizando un solo fotosistema con bacterioclorofila para la fotofosforilación cíclica en la producción de ATP. Sin embargo, también utilizan este mismo fotosistema para la generación de energía reductora, mediante la extracción periódica de electrones a NAD+. El uso de flujo electrónico inverso es innecesario, sin embargo, ya que el portador inicial, la ferredoxina (Fd) tiene un E0' con un potencial de reducción más negativo que el NAD (P). Se requiere un donador de electrones externo, típicamente mediante el uso de H2S o tiosulfato. Así, las bacterias verdes operan como fotoautótrofos, alternando el uso de su fotosistema para ATP o NAD (P) H.
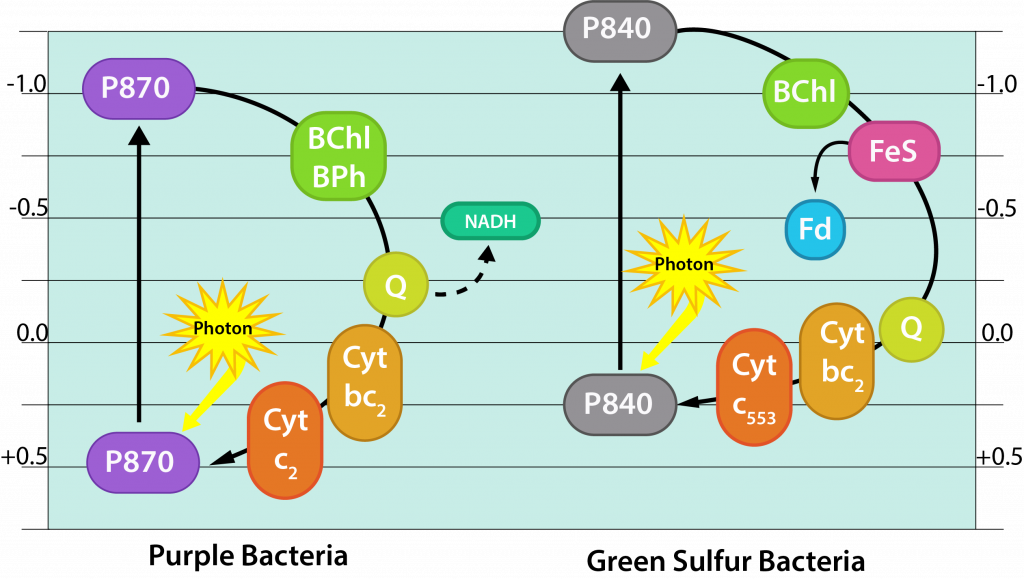
Bacterias Fototróficas Verde y Morada.
Fototrofia Oxigénica
La fototrofia oxigénica es utilizada por cianobacterias que contienen clorofila a, con dos fotosistemas distintos, cada uno con centros de reacción separados. Esto permite la generación tanto de ATP como de potencia reductora en un solo proceso, facilitando el crecimiento fotoautotrófico a través de la fijación de CO2. Esto puede denominarse apropiadamente como fotosíntesis y es el mismo proceso utilizado por las plantas, comúnmente conocido como la “vía Z”.
El proceso comienza cuando la energía lumínica disminuye el potencial de reducción de las moléculas de clorofila a P680 contenidas en el fotosistema II (PSII). Los electrones son luego pasados a través de una cadena de transporte de electrones, generando ATP a través de una fuerza motriz protónica. Luego los electrones pasan al fotosistema I (PSI), donde son golpeados por otro fotón de luz, disminuyendo aún más su potencial de reducción. Luego, los electrones se pasan a través de una cadena de transporte de electrones diferente, eventualmente pasando a NADP+ para la formación de NADPH.
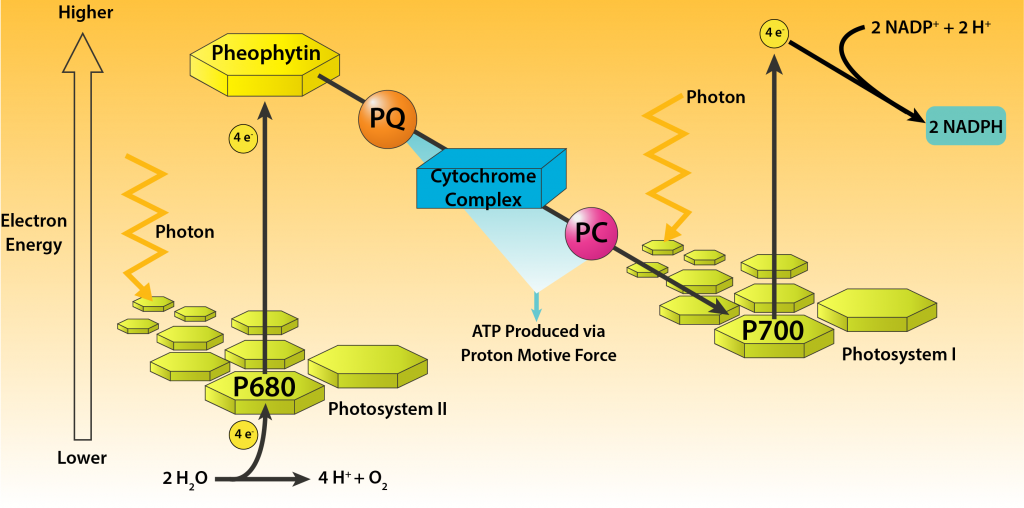
Panorama de la Fotosíntesis Oxígena en Cianobacterias.
El proceso es un ejemplo de fotofosforilación no cíclica, ya que los electrones no son devueltos al fotosistema original. Por lo tanto, se requiere un donador de electrones externo para permitir que el proceso se repita. El agua, que se encuentra en el lado derecho del par redox O2/H2O, es normalmente un donador de electrones pobre, debido a su potencial de reducción extremadamente positivo. Pero el potencial de reducción de la clorofila a P680 es aún más positivo cuando no se excita, permitiendo que el agua sirva como donante de electrones. La hidrólisis del agua conduce a la evolución del oxígeno, un subproducto bienvenido para todos los organismos que utilizan la respiración aeróbica. Se piensa que las cianobacterias son responsables de la oxigenación de la Tierra, permitiendo el desarrollo de la respiración aeróbica como forma de metabolismo.
Existen algunas condiciones bajo las cuales las cianobacterias solo usan PSI, esencialmente realizando una forma de fototrofia anoxígena, a pesar de su posesión de clorofila a. Esto ocurre dentro de los heterocistos de las cianobacterias, donde el oxígeno inactiva las enzimas nitrogenasas. Los heterocistos degradan el PSII, asegurando que el oxígeno no se producirá como subproducto, al tiempo que permiten la producción de ATP con el fotosistema restante.
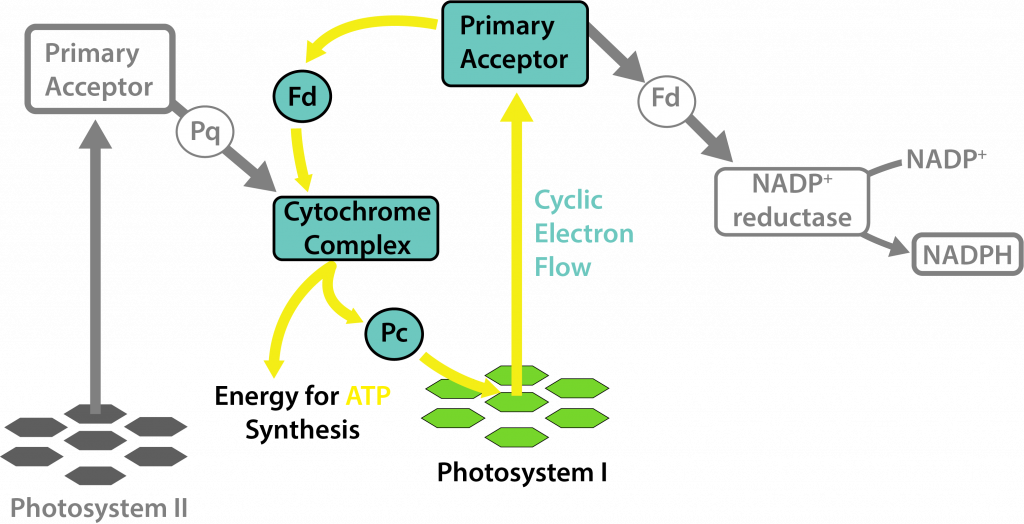
Fototrofia Libre de Oxígeno en Heterocistos Cianobacterianos.
Fototrofia a base de rodopsina
Una forma inusual de fototrofia es utilizada por las arqueas, sin el uso de clorofila o bacterioclorofila. En cambio, estos organismos utilizan una bacteriorodopsina (más apropiadamente llamada arcodopsina), una molécula retiniana relacionada con la que se encuentra en los ojos vertebrados. Cuando la rodopsina absorbe luz sufre un cambio conformacional, bombeando un protón a través de la membrana celular y conduciendo al desarrollo de una fuerza motriz protónica, sin la participación de una cadena de transporte de electrones.
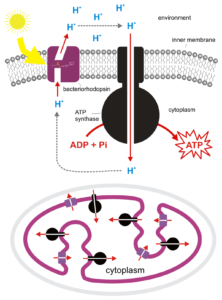
Fototrofia a Base de Rodopsina. Por Darekk2 (Obra propia) [CC BY-SA 3.0], vía Wikimedia Commons
Palabras clave
fototrofia, fotosíntesis, fotoautótrofo, fotoheterótrofo, clorofilas, bacterioclorofilas, carotenoide, ficobiliproteína, ficoeritrina, ficocianina, clorosoma, antenas, centros de reacción, fotofosforilación, fotofosforilación cíclica, fotofosforilación no cíclica, bacterias fototróficas moradas, fototrofia anoxigénica, flujo inverso de electrones, bacterias fototróficas verdes, fototrofia oxigénica, vía Z, fotosistema II (PSII), fotosistema I (PSI), fototrofia basada en rodopsina, bacteriorodopsina/arqueorodopsina.
Preguntas/Objetivos Esenciales
- ¿Cómo difieren la fototrofia y la fotosíntesis? ¿Cómo encajan los fotoautótrofos y fotoheterótrofos con cada término?
- ¿Cuál es la diferencia entre clorofila, bacterioclorofila y pigmentos accesorios de fototrofos?
- ¿Cómo se forma el ATP durante la fotofosforilación? ¿Qué mecanismos se utilizan?
- ¿Qué es la fototrofia anoxígena? ¿En qué se diferencia el proceso entre las bacterias fototróficas moradas y verdes? ¿En qué se diferencia cada proceso entre fotoautótrofos y fotohetertrofos? ¿Qué es el flujo inverso de electrones?
- Ser capaz de trazar y explicar la fotosíntesis cíclica o anoxígena en bacterias fotosintéticas Púrpura y Verde —incluyen pigmentos utilizados, productos formados, fuente de electrones, aceptor final de electrones y cómo se forma ATP.
- Ser capaz de trazar y explicar la vía Z de la fotosíntesis en Cyanobacterium incluyendo pigmento utilizado, productos formados, fuente de electrones, aceptor final de electrones y cómo se forma ATP.
- ¿Qué es la fotosíntesis cíclica en las cianobacterias? ¿Cuándo utiliza este tipo de proceso las cianobacterias?
- ¿Qué es la fototrofia basada en rodopsina? ¿Qué implica y qué organismos lo utilizan? ¿En qué condiciones se utiliza?
Preguntas Exploratorias (OPCIONAL)
- ¿En qué condiciones sería ventajoso que un microbio operara como fotoheterótrofo, a diferencia de fotoautótrofo?