
10.2: Estructura y Función del ADN
- Page ID
- 54500

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Objetivos de aprendizaje
- Describir la estructura bioquímica de los desoxirribonucleótidos
- Identificar los pares de bases utilizados en la síntesis de desoxirribonucleótidos
- Explicar por qué la doble hélice del ADN se describe como antiparalela
En Metabolismo microbiano, se discutieron tres clases de macromoléculas: proteínas, lípidos y carbohidratos. En este capítulo, discutiremos una cuarta clase de macromoléculas: los ácidos nucleicos. Al igual que otras macromoléculas, los ácidos nucleicos están compuestos por monómeros, llamados nucleótidos, que se polimerizan para formar cadenas grandes. Cada cadena de ácido nucleico contiene ciertos nucleótidos que aparecen en un cierto orden dentro de la cadena, llamada su secuencia de bases. La secuencia de bases del ácido desoxirribonucleico (ADN) es responsable de portar y retener la información hereditaria en una célula. En Mecanismos de Genética Microbiana, discutiremos en detalle las formas en que el ADN utiliza su propia secuencia de bases para dirigir su propia síntesis, así como la síntesis de ARN y proteínas, que, a su vez, da lugar a productos con diversa estructura y función. En esta sección, discutiremos la estructura básica y función del ADN.
nucleótidos de ADN
Los bloques de construcción de los ácidos nucleicos son nucleótidos. Los nucleótidos que componen el ADN se denominan desoxirribonucleótidos. Los tres componentes de un desoxirribonucleótido son un azúcar de cinco carbonos llamado desoxirribosa, un grupo fosfato y una base nitrogenada, una estructura de anillo que contiene nitrógeno que es responsable del emparejamiento de bases complementarias entre cadenas de ácido nucleico (Figura\(\PageIndex{1}\)). Los átomos de carbono de la desoxirribosa de cinco carbonos están numerados 1′, 2′, 3′, 4′ y 5′ (1ʹ se lee como “un primo”). Un nucleósido comprende el azúcar de cinco carbonos y la base nitrogenada.
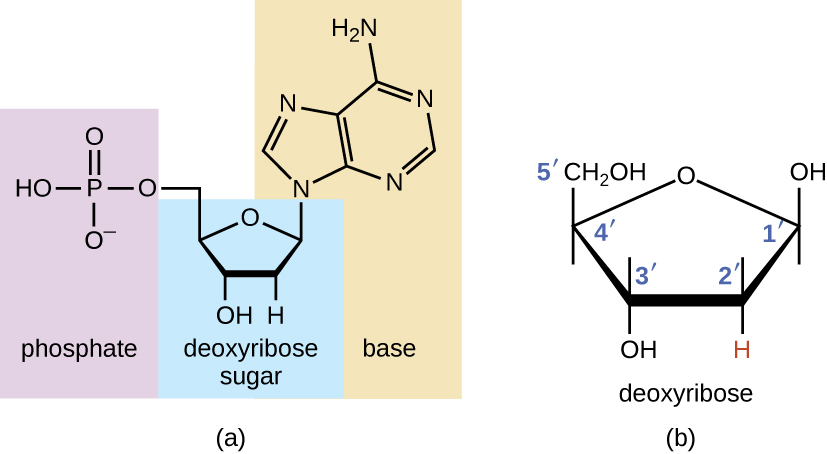
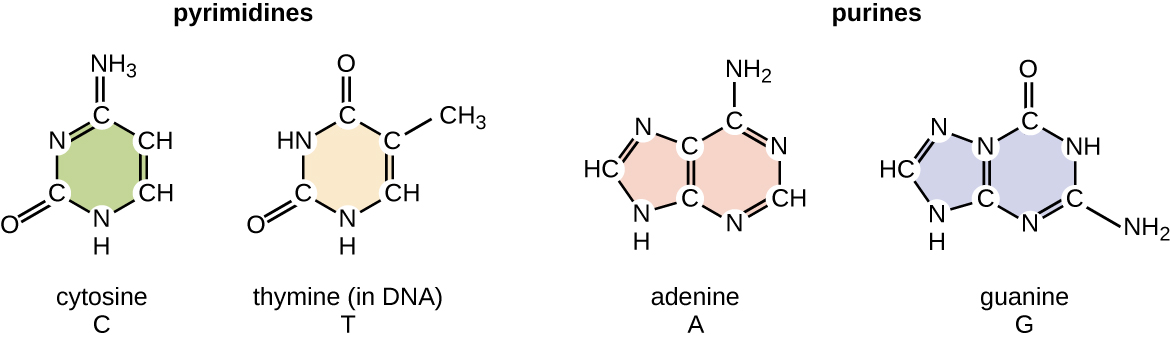
El desoxirribonucleótido se nombra de acuerdo con las bases nitrogenadas (Figura\(\PageIndex{2}\)). Las bases nitrogenadas adenina (A) y guanina (G) son las purinas; tienen una estructura de doble anillo con un anillo de seis carbonos fusionado a un anillo de cinco carbonos. Las pirimidinas, citosina (C) y timina (T), son bases nitrogenadas más pequeñas que solo tienen una estructura de anillo de seis carbonos.
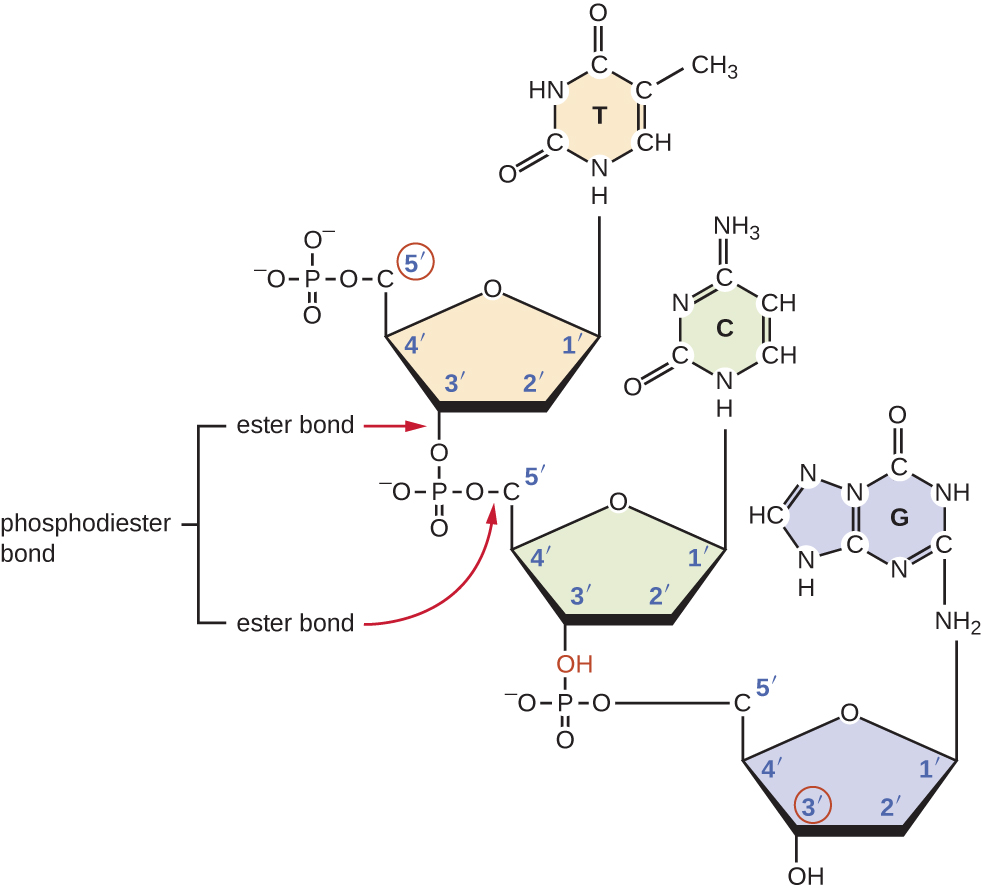
Los nucleósidos trifosfatos individuales se combinan entre sí mediante enlaces covalentes conocidos como enlaces fosfodiéster 5'-3', o enlaces mediante los cuales el grupo fosfato unido al carbono 5' del azúcar de un nucleótido se une al grupo hidroxilo del carbono 3' del azúcar del siguiente nucleótido. La unión fosfodiéster entre nucleótidos forma la cadena principal de azúcar-fosfato, la estructura alternante de azúcar-fosfato que compone el marco de una cadena de ácido nucleico (Figura\(\PageIndex{3}\)). Durante el proceso de polimerización se utilizan desoxinucleótidos trifosfatos (dNTP). Para construir la cadena principal de azúcar-fosfato, los dos fosfatos terminales se liberan del dNTP como pirofosfato. La cadena resultante de ácido nucleico tiene un grupo fosfato libre en el extremo 5' de carbono y un grupo hidroxilo libre en el extremo 3' de carbono. Los dos grupos fosfato no utilizados del nucleótido trifosfato se liberan como pirofosfato durante la formación del enlace fosfodiéster. Posteriormente se hidroliza el pirofosfato, liberando la energía utilizada para impulsar la polimerización de nucleótidos.
Ejercicio\(\PageIndex{1}\)
¿Qué se entiende por los extremos 5' y 3' de una cadena de ácido nucleico?
Descubriendo la Doble Hélice
A principios de la década de 1950, se habían acumulado pruebas considerables que indicaban que el ADN era el material genético de las células, y ahora la carrera estaba en marcha para descubrir su estructura tridimensional. Alrededor de esta época, el bioquímico austriaco Erwin Chargaff 1 (1905—2002) examinó el contenido de ADN en diferentes especies y descubrió que no se encontraron adenina, timina, guanina y citosina en cantidades iguales, y que variaba de especie a especie, pero no entre individuos de la misma especies. Encontró que la cantidad de adenina estuvo muy cerca de igualar la cantidad de timina, y la cantidad de citosina estuvo muy cerca de igualar la cantidad de guanina, o A = T y G = C. Estas relaciones también se conocen como reglas de Chargaff.
Otros científicos también estaban explorando activamente este campo a mediados del siglo XX. En 1952, el científico estadounidense Linus Pauling (1901—1994) fue el principal químico estructural del mundo y el favorito de las probabilidades para resolver la estructura del ADN. Pauling había descubierto anteriormente la estructura de las hélices de proteína α, usando difracción de rayos X, y, basándose en imágenes de difracción de rayos X del ADN hechas en su laboratorio, propuso un modelo de ADN de triple cadena. 2 Al mismo tiempo, los investigadores británicos Rosalind Franklin (1920-1958) y su estudiante de posgrado R.G. Gosling también estaban utilizando difracción de rayos X para comprender la estructura del ADN (Figura\(\PageIndex{4}\)). Fue la experiencia científica de Franklin lo que resultó en la producción de imágenes de difracción de rayos X más bien definidas de ADN que mostrarían claramente la estructura general de doble hélice del ADN.

James Watson (1928—), un científico estadounidense, y Francis Crick (1916-2004), un científico británico, trabajaban juntos en la década de 1950 para descubrir la estructura del ADN. Utilizaron las reglas de Chargaff y las difraccionimágenes de rayos X de Franklin y Wilkins de fibras de ADN para juntar el apareamiento purina-pirimidina de la molécula de ADN de doble hélice (Figura\(\PageIndex{5}\)). En abril de 1953, Watson y Crick publicaron su modelo de la doble hélice de ADN en Nature. 3 El mismo número incluyó además artículos de Wilkins y colegas, 4 así como de Franklin y Gosling, 5 cada uno describiendo diferentes aspectos de la estructura molecular del ADN. En 1962, James Watson, Francis Crick y Maurice Wilkins recibieron el Premio Nobel de Fisiología y Medicina. Desafortunadamente, para entonces Franklin había muerto, y los premios Nobel en ese momento no se otorgaban póstumamente. Sin embargo, continuaron los trabajos sobre el aprendizaje de la estructura del ADN. En 1973, Alexander Rich (1924-2015) y sus colegas pudieron analizar los cristales de ADN para confirmar y dilucidar aún más la estructura del ADN. 6

Ejercicio\(\PageIndex{2}\)
¿A qué científicos se les da la mayor parte del crédito por describir la estructura molecular del ADN?
Estructura del ADN
Watson y Crick propusieron que el ADN está compuesto por dos hebras que se tuercen una alrededor de la otra para formar una hélice diestra. Las dos cadenas de ADN son antiparalelas, de manera que el extremo 3' de una hebra se enfrenta al extremo 5' de la otra (Figura\(\PageIndex{6}\)). El extremo 3' de cada hebra tiene un grupo hidroxilo libre, mientras que el extremo 5' de cada hebra tiene un grupo fosfato libre. El azúcar y el fosfato de los nucleótidos polimerizados forman el esqueleto de la estructura, mientras que las bases nitrogenadas se apilan en su interior. Estas bases nitrogenadas en el interior de la molécula interactúan entre sí, emparejando bases.
El análisis de los patrones de difracción del ADN ha determinado que hay aproximadamente 10 bases por giro en el ADN. El espaciamiento asimétrico de las cadenas principales de azúcar-fosfato genera surcos mayores (donde la columna vertebral está muy separada) y surcos menores (donde la columna vertebral está muy próxima) (Figura\(\PageIndex{6}\)). Estos surcos son localizaciones donde las proteínas pueden unirse al ADN. La unión de estas proteínas puede alterar la estructura del ADN, regular la replicación o regular la transcripción del ADN en ARN.
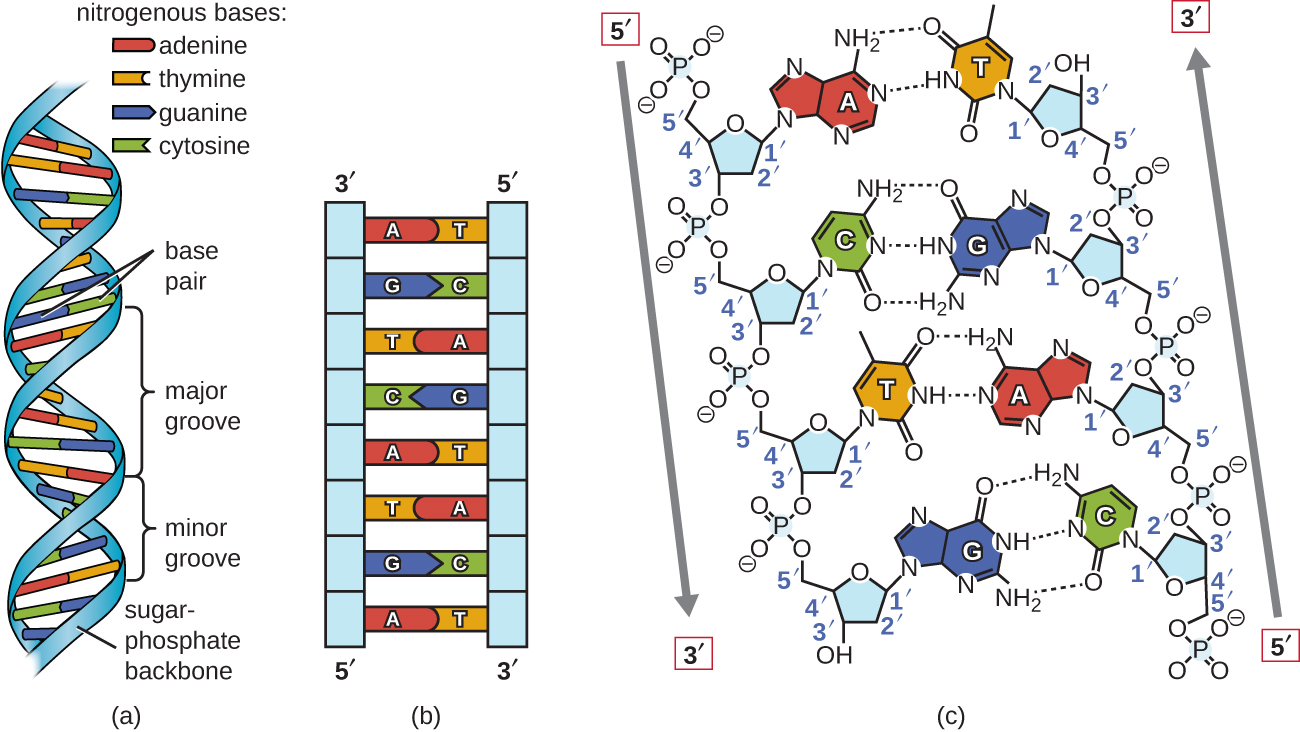
El emparejamiento de bases se lleva a cabo entre una purina y una pirimidina. En el ADN, la adenina (A) y la timina (T) son pares de bases complementarios, y la citosina (C) y la guanina (G) también son pares de bases complementarias, explicando las reglas de Chargaff (Figura\(\PageIndex{7}\)). Los pares de bases se estabilizan mediante enlaces de hidrógeno; la adenina y la timina forman dos enlaces de hidrógeno entre ellos, mientras que la citosina y la guanina forman tres enlaces de hidrógeno entre ellos.
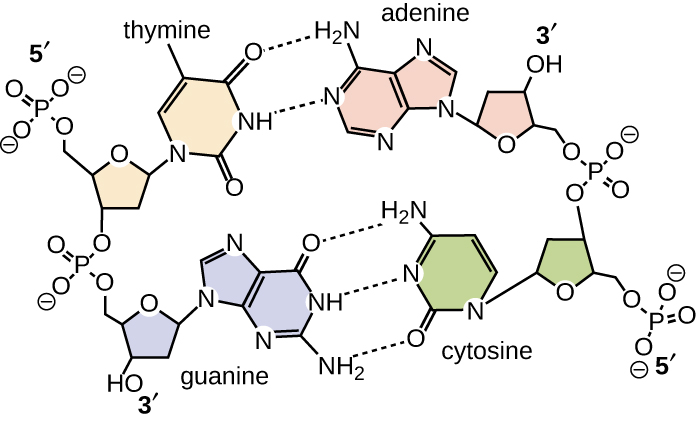
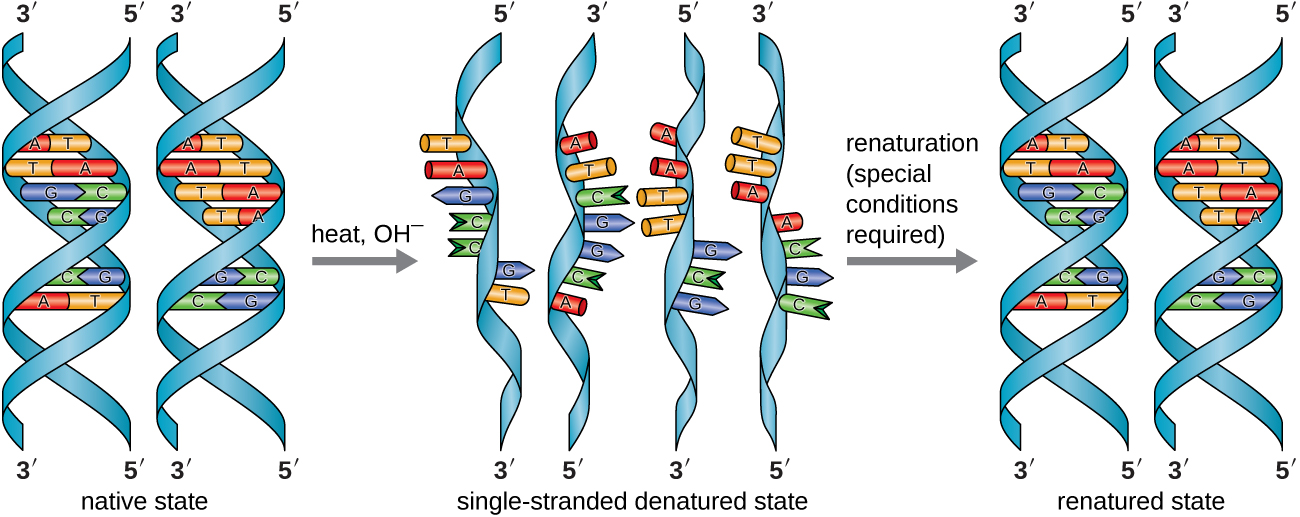
En el laboratorio, exponer las dos cadenas de ADN de la doble hélice a altas temperaturas o a ciertos productos químicos puede romper los enlaces de hidrógeno entre bases complementarias, separando así las cadenas en dos cadenas individuales separadas de ADN (ADN monocatenario [ADNss]). Este proceso se denomina desnaturalización del ADN y es análogo a la desnaturalización de proteínas, como se describe en Proteínas. Las cadenas de ssDNA también se pueden volver a unir como ADN bicatenario (dsDNA), mediante reasociación o renaturalización enfriando o eliminando los desnaturalizantes químicos, permitiendo que estos enlaces de hidrógeno se reformen. La capacidad de manipular artificialmente el ADN de esta manera es la base de varias técnicas importantes en biotecnología (Figura\(\PageIndex{8}\)). Debido al enlace de hidrógeno adicional entre el par de bases C = G, el ADN con un alto contenido de GC es más difícil de desnaturalizar que el ADN con un menor contenido de GC.
Vea una animación sobre la estructura del ADN desde el Centro de Aprendizaje de ADN para obtener más información.
Ejercicio\(\PageIndex{3}\)
¿Cuáles son los dos pares de bases complementarias del ADN y cómo se unen?
Función ADN
El ADN almacena la información necesaria para construir y controlar la célula. La transmisión de esta información de células madre a hija se denomina transferencia génica vertical y se produce a través del proceso de replicación del ADN. El ADN se replica cuando una célula hace una copia duplicada de su ADN, luego la célula se divide, dando como resultado la distribución correcta de una copia de ADN a cada célula resultante. El ADN también puede degradarse enzimáticamente y usarse como fuente de nucleósidos y nucleótidos para la célula. A diferencia de otras macromoléculas, el ADN no cumple un papel estructural en las células.
Ejercicio\(\PageIndex{4}\)
¿Cómo transmite el ADN la información genética a la descendencia?
Preparando el camino para las mujeres en las profesiones científicas y de la salud
Históricamente, las mujeres han estado subrepresentadas en las ciencias y en la medicina, y muchas veces sus contribuciones pioneras han pasado relativamente desapercibidas. Por ejemplo, aunque Rosalind Franklin realizó los estudios de difracción de rayos X demostrando la doble estructura helicoidal del ADN, son Watson y Crick quienes se hicieron famosos por este descubrimiento, basándose en sus datos. Aún queda una gran controversia sobre si su adquisición de sus datos fue apropiada y si los conflictos de personalidad y el sesgo de género contribuyeron al retraso en el reconocimiento de sus contribuciones significativas. De igual manera, Barbara McClintock realizó un trabajo pionero en genética del maíz (maíz) desde los años treinta hasta los 50, descubriendo transposones (genes saltarines), pero no fue reconocida hasta mucho después, recibiendo el Premio Nobel de Fisiología o Medicina en 1983 (Figura\(\PageIndex{9}\)).
Hoy en día, las mujeres siguen estando subrepresentadas en muchos campos de la ciencia y la medicina. Si bien más de la mitad de las licenciaturas en ciencias se otorgan a mujeres, sólo 46% de los doctorados en ciencias se otorgan a mujeres. En la academia, el número de mujeres en cada nivel de avance profesional sigue disminuyendo, con mujeres ocupando menos de un tercio de los puestos de científicos de nivel doctorado en puestos de seguimiento de tenencia, y menos de una cuarta parte de las cátedras completas en colegios y universidades de 4 años. 7 Incluso en las profesiones de la salud, como casi todos los demás campos, las mujeres suelen estar subrepresentadas en muchas carreras médicas y ganan significativamente menos que sus homólogos masculinos, como se muestra en un estudio de 2013 publicado por el Journal of the American Medical Association. 8
¿Por qué siguen existiendo esas disparidades y cómo rompemos estos ciclos? La situación es compleja y probablemente resulta de la combinación de diversos factores, entre ellos cómo la sociedad condiciona los comportamientos de las niñas desde una edad temprana y apoya sus intereses, tanto profesional como personalmente. Algunos han sugerido que las mujeres no pertenecen al laboratorio, entre ellas el ganador del Premio Nobel Tim Hunt, cuyos comentarios públicos de 2015 que sugieren que las mujeres son demasiado emocionales para la ciencia 9 fueron recibidos con una condena generalizada.
Quizás las niñas deberían ser apoyadas más desde una edad temprana en las áreas de ciencias y matemáticas (Figura\(\PageIndex{9}\)). Los programas de ciencia, tecnología, ingeniería y matemáticas (STEM) patrocinados por la Asociación Americana de Mujeres Universitarias (AAUW) 10 y la Administración Nacional de Aeronáutica y del Espacio (NASA) 11 son excelentes ejemplos de programas que ofrecen dicho apoyo. Las contribuciones de las mujeres en la ciencia deben darse a conocer más ampliamente entre el público, y el marketing dirigido a las niñas debería incluir más imágenes de científicas y profesionales médicas de éxito histórico y profesional, alentando a todas las mentes jóvenes brillantes, incluidas niñas y mujeres, a perseguir carreras en ciencias y medicina.

Enfoque Clínico: Parte 2
Con base en sus síntomas, el médico de Alex sospecha que padece una enfermedad transmitida por los alimentos que adquirió durante sus viajes. Las posibilidades incluyen infección bacteriana (por ejemplo, E. coli enterotoxigénica, Vibrio cholerae, Campylobacter jejuni, Salmonella), infección viral (rotavirus o norovirus) o infección por protozoos (Giardia lamblia, Cryptosporidium parvum , o Entamoeba histolytica).
Su médico ordena una muestra de heces para identificar posibles agentes causantes (por ejemplo, bacterias, quistes) y buscar la presencia de sangre porque ciertos tipos de agentes infecciosos (como C. jejuni, Salmonella y E. histolytica) están asociados con la producción de sangre taburetes.
La muestra de heces de Alex no mostró sangre ni quistes. Después del análisis de su muestra de heces y con base en su reciente historial de viajes, el médico del hospital sospechó que Alex padecía diarrea del viajero causada por E. coli enterotoxigénica (ETEC), el agente causante de la mayoría de las diarreas del viajero. Para verificar el diagnóstico y descartar otras posibilidades, el médico de Alex ordenó una prueba diagnóstica de laboratorio de su muestra de heces para buscar secuencias de ADN que codificaran factores de virulencia específicos de ETEC. El médico le indicó a Alex que tomara muchos líquidos para reemplazar lo que estaba perdiendo y lo dio de alta del hospital.
ETEC produce varios factores de virulencia codificados por plásmidos que la hacen patógena en comparación con E. coli típica. Estas incluyen las toxinas secretadas enterotoxina lábil al calor (LT) y enterotoxina termoestable (ST), así como factor de colonización (CF). Tanto el LT como el ST provocan la excreción de iones cloruro de las células intestinales a la luz intestinal, provocando una consecuente pérdida de agua de las células intestinales, resultando en diarrea. La CF codifica una proteína bacteriana que ayuda a permitir que la bacteria se adhiera al revestimiento del intestino delgado.
Ejercicio\(\PageIndex{5}\)
¿Por qué el médico de Alex utilizó el análisis genético en lugar del aislamiento de bacterias de la muestra de heces o la tinción directa de Gram de la muestra de heces sola?
Conceptos clave y resumen
- Los ácidos nucleicos están compuestos por nucleótidos, cada uno de los cuales contiene un azúcar pentosa, un grupo fosfato y una base nitrogenada. Los desoxirribonucleótidos dentro del ADN contienen desoxirribosa como el azúcar pentosa.
- El ADN contiene las pirimidinas citosina y timina, y las purinas adenina y guanina.
- Los nucleótidos están unidos entre sí por enlaces fosfodiéster entre el grupo fosfato 5' de un nucleótido y el grupo hidroxilo 3' de otro. Una cadena de ácido nucleico tiene un grupo fosfato libre en el extremo 5' y un grupo hidroxilo libre en el extremo 3'.
- Chargaff descubrió que la cantidad de adenina es aproximadamente igual a la cantidad de timina en el ADN, y que la cantidad de guanina es aproximadamente igual a la citosina. Posteriormente se determinó que estas relaciones se debían al emparejamiento de bases complementarias.
- Watson y Crick, basándose en el trabajo de Chargaff, Franklin y Gosling, y Wilkins, propusieron el modelo de doble hélice y el emparejamiento de bases para la estructura del ADN.
- El ADN está compuesto por dos cadenas complementarias orientadas antiparalelas entre sí con las cadenas principales de fosfodiéster en el exterior de la molécula. Las bases nitrogenadas de cada hebra se enfrentan entre sí y las bases complementarias se unen entre sí, estabilizando la doble hélice.
- El calor o los químicos pueden romper los enlaces de hidrógeno entre bases complementarias, desnaturalizando el ADN. Enfriar o eliminar químicos puede conducir a renaturalización o reasociación del ADN al permitir que los enlaces de hidrógeno se reformen entre bases complementarias.
- El ADN almacena las instrucciones necesarias para construir y controlar la célula. Esta información se transmite de padres a hijos a través de la transferencia vertical de genes.
Notas al pie
- 1 N. Kresge et al. “Reglas de Chargaff: La obra de Erwin Chargaff”. Revista de Química Biológica 280 (2005) :e21.
- 2 L. Pauling, “Una estructura propuesta para los ácidos nucleicos”. Actas de la Academia Nacional de Ciencias de los Estados Unidos de América 39 núm. 2 (1953) :84—97.
- 3 J.D. Watson, F.H.C. Crick. “Una estructura para ácido nucleico desoxirribosa”. Naturaleza 171 núm. 4356 (1953) :737—738.
- 4 M.H.F. Wilkins et al. “Estructura Molecular de los Ácidos Nucleicos de Desoxipentosa.” Naturaleza 171 núm. 4356 (1953) :738—740.
- 5 R. Franklin, R.G. Gosling. “Configuración Molecular en Timonucleato de Sodio”. Naturaleza 171 núm. 4356 (1953) :740—741.
- 6 R.O. Day et al. “Un Fragmento Cristalino de la Doble Hélice: La Estructura del Dinucleósido Fosfato Guanilyl-3',5'-citidina”. Actas de la Academia Nacional de Ciencias de los Estados Unidos de América 70 núm. 3 (1973) :849—853.
- 7 N.H. Wolfinger “Para las científicas, no hay buen momento para tener hijos”. El Atlántico 29 de julio de 2013. www.theatlantic.com/sexes/arc... ildren/278165/.
- 8 S.A. Seabury et al. “Tendencias en las ganancias de los profesionales de la salud masculinos y femeninos en Estados Unidos, 1987 a 2010”. Revista de la Asociación Médica Americana Medicina Interna 173 núm. 18 (2013) :1748—1750.
- 9 E. Chung. “Tim Hunt, sexismo y ciencia: El verdadero 'problema con las chicas en los laboratorios”. Noticias CBC Tecnología y Ciencia, 12 de junio de 2015. http://www.cbc.ca/news/technology/ti...labs-1.3110133. Accedido 04/08/2016.
- 10 Asociación Americana de Mujeres Universitarias. “Construyendo un Pipeloducto STEM para Niñas y Mujeres”. www.aauw.org/what-we-do/stem-education/. Consultado el 10 de junio de 2016.
- 11 Administración Nacional de Aeronáutica y del Espacio. “Programas de Divulgación: Iniciativa Mujeres y Niñas”. http://women.nasa.gov/outreach-programs/. Consultado el 10 de junio de 2016.