
17.3: Defensas celulares
- Page ID
- 54366

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Objetivos de aprendizaje
- Identificar y describir los componentes de la sangre
- Explicar el proceso por el cual se forman los elementos formados de la sangre (hematopoyesis)
- Describir las características de los elementos formados que se encuentran en la sangre periférica, así como sus respectivas funciones dentro del sistema inmune innato
En la sección anterior, se discutieron algunos de los mediadores químicos que se encuentran en el plasma, la porción líquida de la sangre. La porción no fluida de la sangre consiste en varios tipos de elementos formados, llamados así porque todos están formados a partir de las mismas células madre que se encuentran en la médula ósea. Las tres categorías principales de elementos formados son: glóbulos rojos (RBC), también llamados eritrocitos; plaquetas, también llamadas trombocitos; y glóbulos blancos (WBC), también llamados leucocitos.
Los glóbulos rojos son los principales responsables de transportar oxígeno a los tejidos. Las plaquetas son fragmentos celulares que participan en la formación de coágulos sanguíneos y la reparación de tejidos. Varios tipos diferentes de WBC participan en diversos mecanismos inespecíficos de inmunidad innata y adaptativa. En esta sección, nos centraremos principalmente en los mecanismos innatos de diversos tipos de WBC.
Hematopoyesis
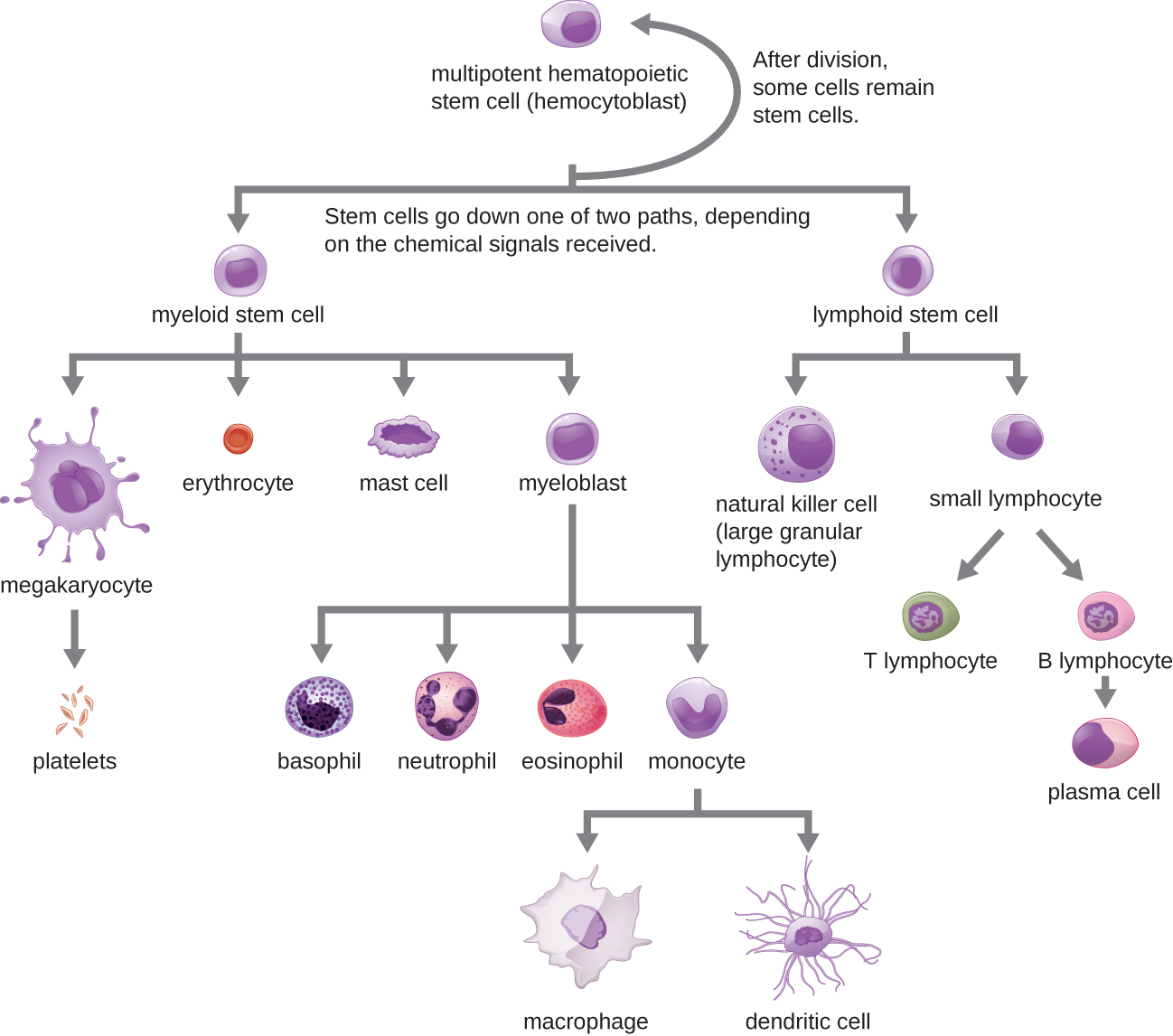
Todos los elementos formados de la sangre se derivan de células madre hematopoyéticas pluripotentes (CMH) en la médula ósea. A medida que las CMH hacen copias de sí mismas en la médula ósea, las células individuales reciben diferentes señales del cuerpo que controlan cómo se desarrollan y maduran. Como resultado, las CMH se diferencian en diferentes tipos de células sanguíneas que, una vez maduras, circulan en sangre periférica. Este proceso de diferenciación, denominado hematopoyesis, se muestra con más detalle en la Figura\(\PageIndex{1}\).
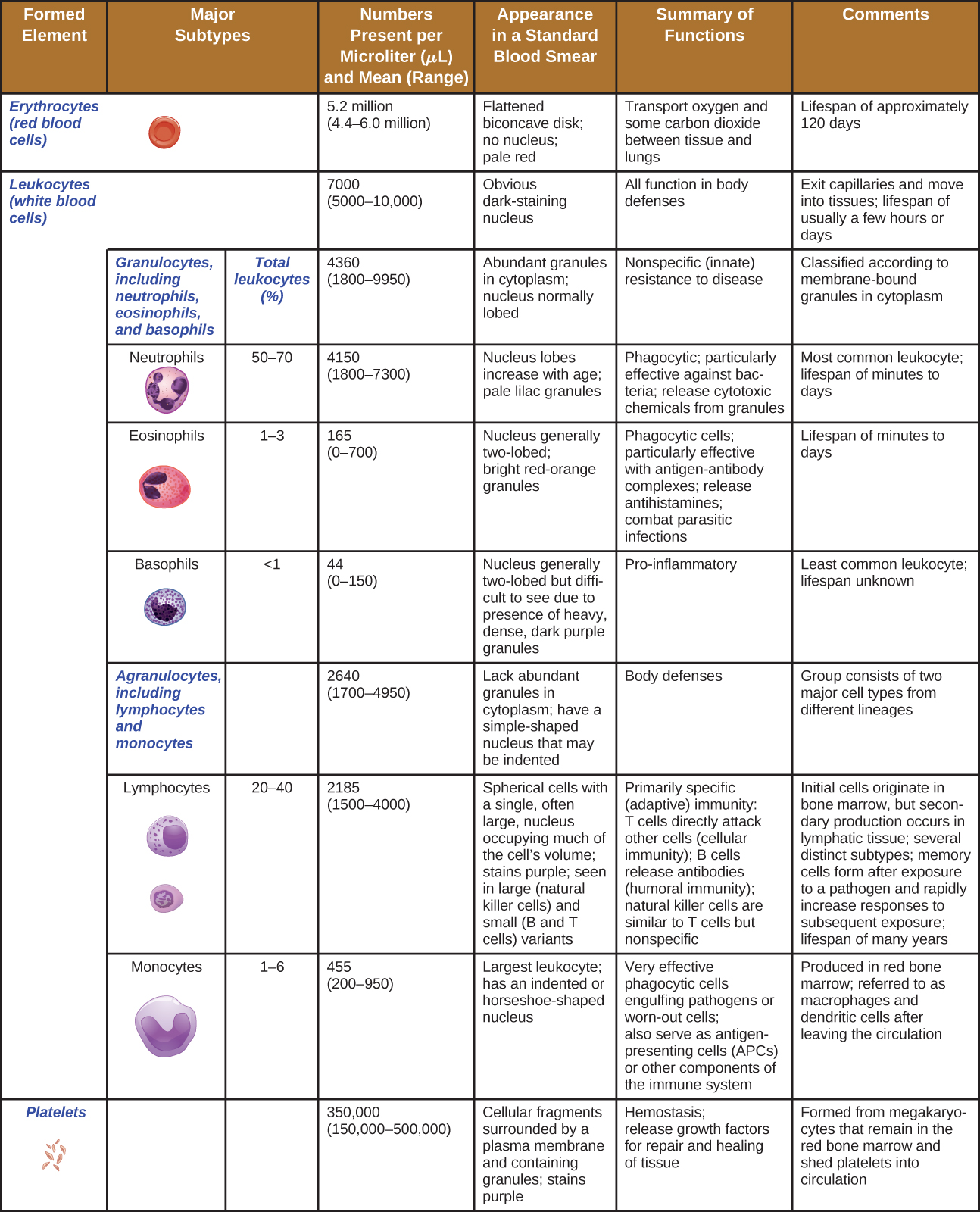
En términos de números puros, la gran mayoría de las CMH se convierten en eritrocitos. Números mucho más pequeños se convierten en leucocitos y plaquetas. Los leucocitos se pueden subdividir en granulocitos, que se caracterizan por numerosos gránulos visibles en el citoplasma, y agranulocitos, que carecen de gránulos. La figura\(\PageIndex{2}\) proporciona una visión general de los diversos tipos de elementos formados, incluyendo sus números relativos, función primaria y esperanza de vida.
Granulocitos
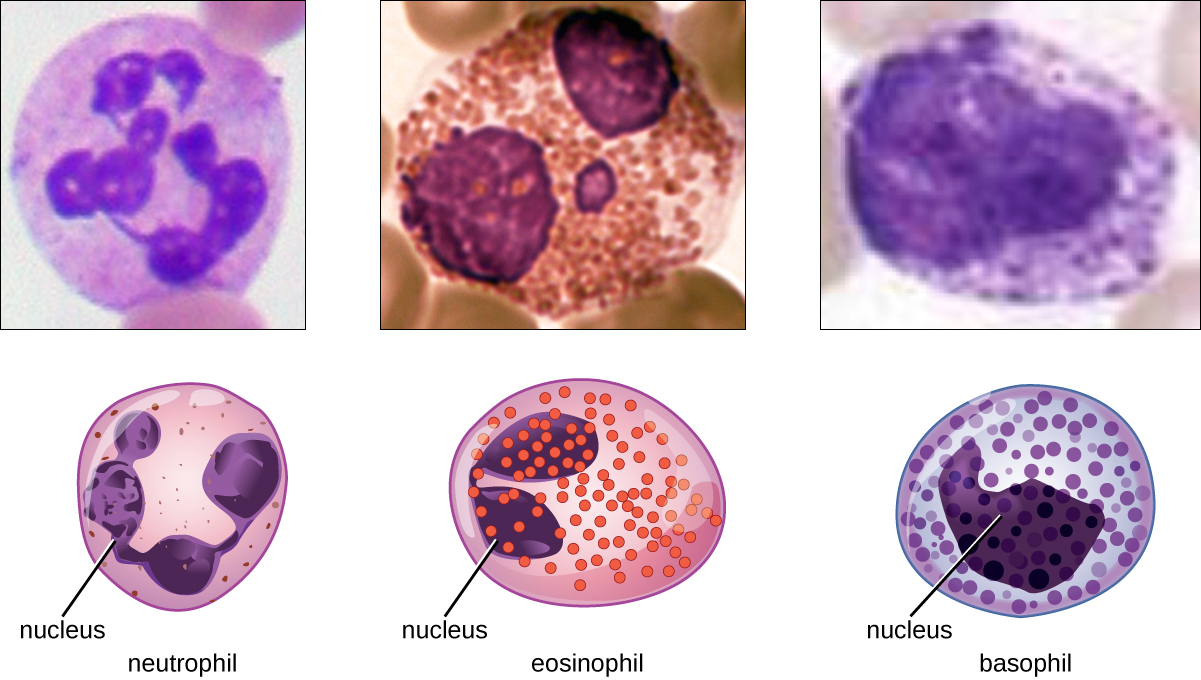
Los diversos tipos de granulocitos se pueden distinguir entre sí en un frotis de sangre por la aparición de sus núcleos y el contenido de sus gránulos, que confieren diferentes rasgos, funciones y propiedades de tinción. Los neutrófilos, también llamados neutrófilos polimorfonucleares (PMN), tienen un núcleo con tres a cinco lóbulos y pequeños, numerosos, gránulos de color lila. Cada lóbulo del núcleo está conectado por una delgada hebra de material a los otros lóbulos. Los eosinófilos tienen menos lóbulos en el núcleo (típicamente 2—3) y gránulos más grandes que tiñen de color naranja rojizo. Los basófilos tienen un núcleo de dos lóbulos y gránulos grandes que se tiñen de azul oscuro o púrpura (Figura\(\PageIndex{3}\)).
Neutrófilos (PMN)
Los neutrófilos (PMN) están frecuentemente involucrados en la eliminación y destrucción de bacterias extracelulares. Son capaces de migrar a través de las paredes de los vasos sanguíneos a zonas de infección bacteriana y daño tisular, donde buscan y matan bacterias infecciosas. Los gránulos de PMN contienen una variedad de defensinas y enzimas hidrolíticas que les ayudan a destruir las bacterias a través de la fagocitosis (descrita con más detalle en Reconocimiento de patógenos y fagocitosis) Además, cuando muchos neutrófilos son traídos a una zona infectada, pueden ser estimulados para liberar moléculas en el tejido circundante para limpiar mejor los agentes infecciosos. Esto se llama desgranulación.
Otro mecanismo utilizado por los neutrófilos son las trampas extracelulares de neutrófilos (NET), que son mallas extruidas de cromatina que están estrechamente asociadas con proteínas y componentes granulados antimicrobianos. La cromatina es ADN con proteínas asociadas (generalmente proteínas histonas, alrededor de las cuales el ADN se envuelve para organizarse y empaquetarse dentro de una célula). Al crear y liberar una estructura de malla o rejilla de cromatina que se acopla con proteínas antimicrobianas, los neutrófilos pueden montar un ataque altamente concentrado y eficiente contra patógenos cercanos. Las proteínas frecuentemente asociadas con TNE incluyen lactoferrina, gelatinasa, catepsina G y mieloperoxidasa. Cada uno tiene un medio diferente para promover la actividad antimicrobiana, ayudando a los neutrófilos a eliminar patógenos. Las proteínas tóxicas en los TNE pueden matar algunas de las propias células del cuerpo junto con los patógenos invasores. Sin embargo, este daño colateral puede repararse después de que se haya eliminado el peligro de la infección.
A medida que los neutrófilos combaten una infección, se puede observar una acumulación visible de leucocitos, restos celulares y bacterias en el sitio de la infección. Esta acumulación es lo que llamamos pus (también conocido como secreción o drenaje purulento o supurativo). La presencia de pus es una señal de que las defensas inmunitarias se han activado contra una infección; históricamente, algunos médicos creían que inducir la formación de pus en realidad podría promover la curación de heridas. La práctica de promover el “pus loable” (por ejemplo, envolviendo una herida en lana grasienta empapada en vino) se remonta al antiguo médico Galeno en el siglo II d.C., y se practicó en formas variantes hasta el siglo XVII (aunque no fue universalmente aceptada). Hoy en día, este método ya no se practica porque ahora sabemos que no es efectivo. Aunque una pequeña cantidad de formación de pus puede indicar una fuerte respuesta inmune, inducir artificialmente la formación de pus no promueve la recuperación.
Eosinófilos
Los eosinófilos son granulocitos que protegen contra protozoos y helmintos; también juegan un papel en las reacciones alérgicas. Los gránulos de eosinófilos, que absorben fácilmente el colorante ácido rojizo eosina, contienen histamina, enzimas degradativas y un compuesto conocido como proteína básica principal (MBP) (Figura\(\PageIndex{3}\)). La MBP se une a los carbohidratos superficiales de los parásitos, y esta unión se asocia con la alteración de la membrana celular y la permeabilidad de la membrana.
Basófilos
Los basófilos tienen gránulos citoplásmicos de tamaño variado y se nombran por la capacidad de sus gránulos para absorber el colorante básico azul de metileno (Figura\(\PageIndex{3}\)). Su estimulación y desgranulación pueden ser el resultado de múltiples eventos desencadenantes. Los fragmentos de complemento activados C3a y C5a, producidos en las cascadas de activación de las proteínas del complemento, actúan como anafilatoxinas induciendo la desgranulación de basófilos y respuestas inflamatorias. Este tipo de células es importante en las reacciones alérgicas y otras respuestas que involucran inflamación. Uno de los componentes más abundantes de los gránulos de basófilos es la histamina, que se libera junto con otros factores químicos cuando se estimula el basófilo. Estos químicos pueden ser quimiotácticos y pueden ayudar a abrir las brechas entre las células en los vasos sanguíneos. Otros mecanismos para desencadenar basófilos requieren la asistencia de anticuerpos, como se discute en Linfocitos B e Inmunidad Humoral.
Mastocitos
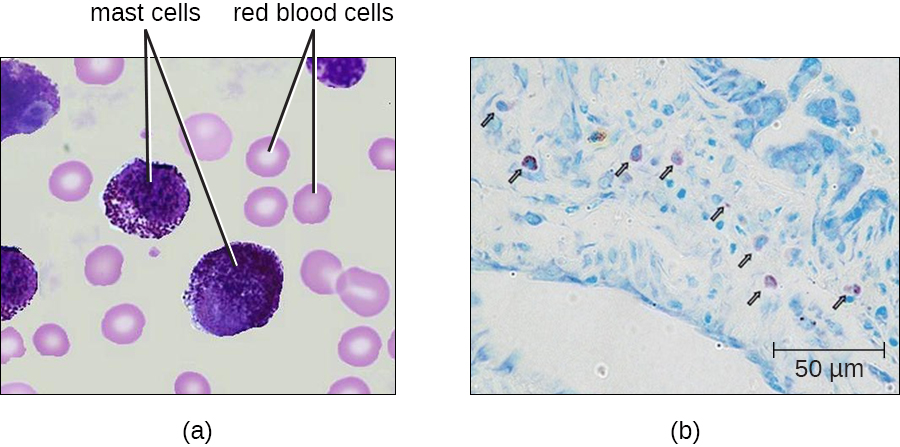
La hematopoyesis también da lugar a mastocitos, que parecen derivarse de la misma célula progenitora mieloide común que los neutrófilos, eosinófilos y basófilos. Funcionalmente, los mastocitos son muy similares a los basófilos, contienen muchos de los mismos componentes en sus gránulos (por ejemplo, histamina) y desempeñan un papel similar en las respuestas alérgicas y otras reacciones inflamatorias. Sin embargo, a diferencia de los basófilos, los mastocitos abandonan la sangre circulante y se encuentran con mayor frecuencia residiendo en los tejidos. A menudo se asocian con vasos sanguíneos y nervios o se encuentran cerca de superficies que interactúan con el ambiente externo, como la piel y las membranas mucosas en diversas regiones del cuerpo (Figura\(\PageIndex{4}\)).
Ejercicio\(\PageIndex{1}\)
- Describir los gránulos y núcleos de neutrófilos, eosinófilos, basófilos y mastocitos.
- Nombrar tres mecanismos antimicrobianos de neutrófilos
Enfoque Clínico: Parte 3
Las pruebas de Angela son negativas para todos los alérgenos comunes, y sus muestras de esputo no contienen presencia anormal de microbios patógenos ni niveles elevados de miembros de la microbiota respiratoria normal. Sin embargo, tiene niveles elevados de citocinas inflamatorias en su sangre.
La hinchazón de su vía aérea aún no ha respondido al tratamiento con antihistamínicos o corticosteroides. Los análisis de sangre adicionales muestran que Angela tiene un recuento de glóbulos blancos levemente elevado pero niveles normales de anticuerpos. Además, tiene un nivel inferior a lo normal de la proteína del complemento C4.
Ejercicio\(\PageIndex{2}\)
- ¿Qué revela esta nueva información sobre la causa de las vías respiratorias constreñidas de Ángela?
- ¿Cuáles son algunas condiciones posibles que podrían conducir a niveles bajos de proteínas del complemento?
Agranulocitos
Como su nombre indica, los agranulocitos carecen de gránulos visibles en el citoplasma. Los agranulocitos pueden clasificarse como linfocitos o monocitos (Figura\(\PageIndex{2}\)). Entre los linfocitos se encuentran las células asesinas naturales, que desempeñan un papel importante en las defensas inmunitarias innatas inespecíficas. Los linfocitos también incluyen las células B y las células T, las cuales se discuten en el siguiente capítulo porque son actores centrales en las defensas inmunitarias adaptativas específicas. Los monocitos se diferencian en macrófagos y células dendríticas, las cuales se denominan colectivamente el sistema de fagocitos mononucleares.
Células asesinas naturales
La mayoría de los linfocitos están involucrados principalmente en la respuesta inmune adaptativa específica, por lo que se discutirá en el siguiente capítulo. Una excepción son las células asesinas naturales (células NK); estos linfocitos mononucleares utilizan mecanismos inespecíficos para reconocer y destruir células que son anormales de alguna manera. Las células cancerosas y las células infectadas con virus son dos ejemplos de anomalías celulares a las que se dirigen las células NK. El reconocimiento de tales células implica un complejo proceso de identificación de marcadores moleculares inhibitorios y activadores en la superficie de la célula diana. Los marcadores moleculares que componen el complejo mayor de histocompatibilidad (MHC) son expresados por células sanas como una indicación de “auto”. Esto se tratará con más detalle en el próximo capítulo. Las células NK son capaces de reconocer marcadores MHC normales en la superficie de las células sanas, y estos marcadores del MHC sirven como una señal inhibidora que previene la activación de las células NK. Sin embargo, las células cancerosas y las células infectadas por virus disminuyen o eliminan activamente la expresión de marcadores del MHC en su superficie. Cuando estos marcadores del MHC están disminuidos o ausentes, la célula NK interpreta esto como una anomalía y una célula en peligro. Esta es una parte del proceso de activación de células NK (Figura\(\PageIndex{5}\)). Las células NK también se activan uniéndose a moléculas moleculares activadoras en la célula diana. Estas moléculas moleculares activadoras incluyen moléculas “auto alteradas” o “no propias”. Cuando una célula NK reconoce una disminución en las moléculas MHC normales inhibitorias y un aumento en las moléculas activadoras en la superficie de una célula, la célula NK se activará para eliminar la célula en peligro.
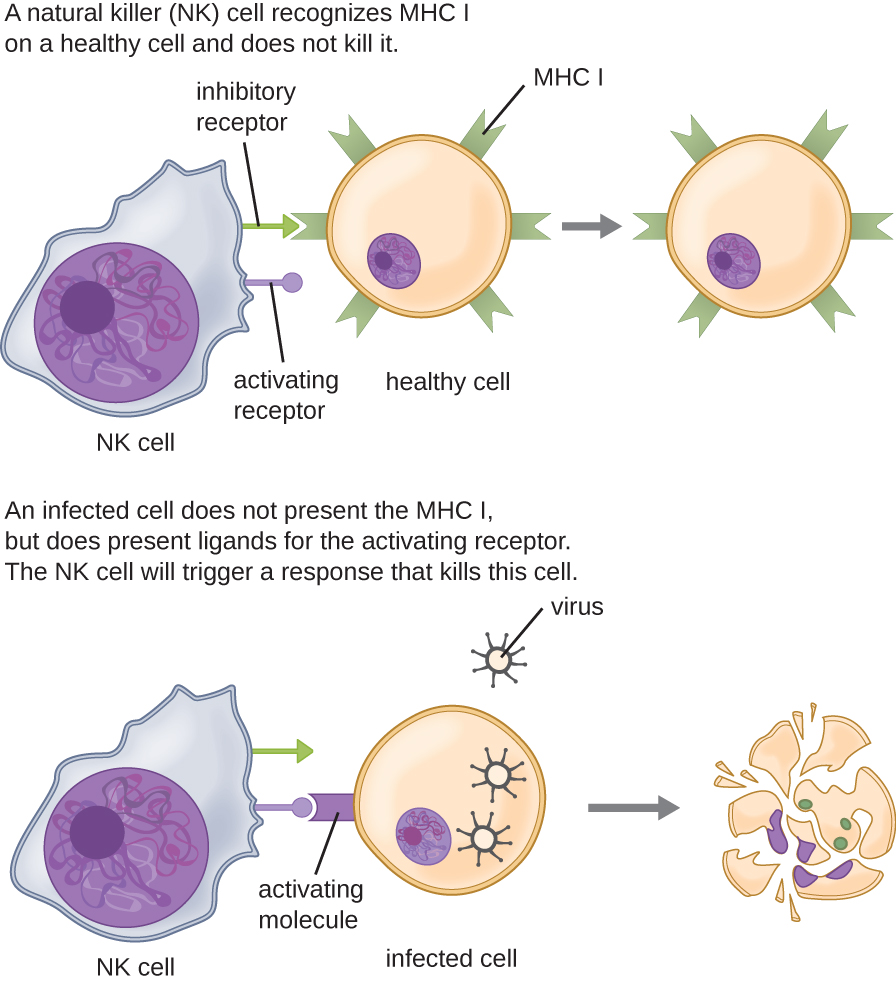
Una vez que una célula ha sido reconocida como diana, la célula NK puede usar varios mecanismos diferentes para matar a su diana. Por ejemplo, puede expresar proteínas de membrana citotóxicas y citocinas que estimulan a la célula diana para que sufra apoptosis, o suicidio celular controlado. Las células NK también pueden usar citotoxicidad mediada por perforina para inducir apoptosis en células diana. Este mecanismo se basa en dos toxinas liberadas de los gránulos en el citoplasma de la célula NK: perforina, una proteína que crea poros en la célula diana, y granzimas, proteasas que ingresan a través de los poros al citoplasma de la célula diana, donde desencadenan una cascada de activación proteica que conduce a la apoptosis. La célula NK se une a la célula diana anormal, libera su carga útil destructiva y se separa de la célula diana. Mientras que la célula diana sufre apoptosis, la célula NK sintetiza más perforina y proteasas para usar en su siguiente diana.
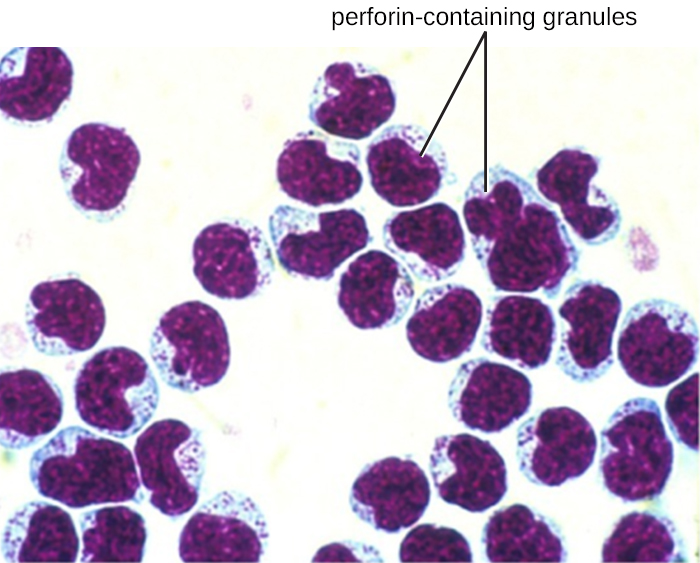
Las células NK contienen estos compuestos tóxicos en gránulos en su citoplasma. Cuando se tiñen, los gránulos son azurófilos y pueden visualizarse bajo un microscopio óptico (Figura\(\PageIndex{6}\)). A pesar de que tienen gránulos, las células NK no se consideran granulocitos porque sus gránulos son mucho menos numerosos que los que se encuentran en los granulocitos verdaderos. Además, las células NK tienen un linaje diferente al de los granulocitos, surgiendo de células madre linfoides en lugar de mieloides (Figura\(\PageIndex{1}\)).
Monocitos
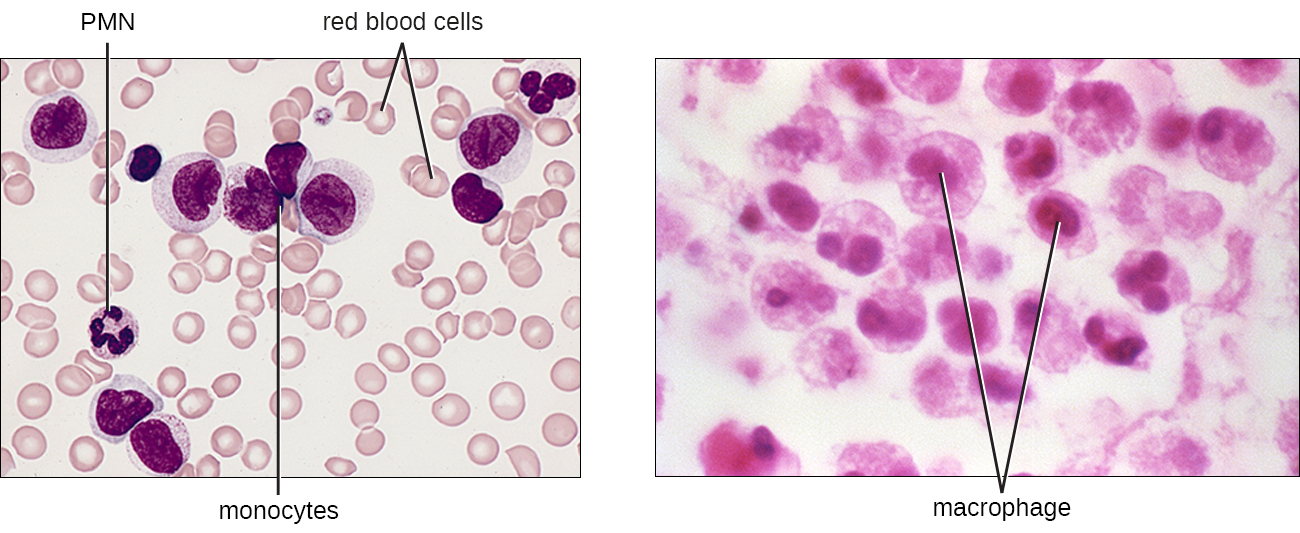
El mayor de los glóbulos blancos, los monocitos tienen un núcleo que carece de lóbulos, y también carecen de gránulos en el citoplasma (Figura\(\PageIndex{7}\)). Sin embargo, son fagocitos efectivos, envolviendo patógenos y células apoptóticas para ayudar a combatir la infección.
Cuando los monocitos salen del torrente sanguíneo y entran en un tejido corporal específico, se diferencian en fagocitos específicos de tejido llamados macrófagos y células dendríticas. Son residentes particularmente importantes del tejido linfoide, así como de sitios y órganos no linfoides. Los macrófagos y las células dendríticas pueden residir en los tejidos corporales durante períodos de tiempo significativos. Los macrófagos en tejidos específicos del cuerpo desarrollan características adecuadas para el tejido particular. No sólo proporcionan protección inmune para el tejido en el que residen sino que también apoyan la función normal de sus células tisulares vecinas a través de la producción de citocinas. A los macrófagos se les dan nombres específicos de tejido, y algunos ejemplos de macrófagos específicos de tejido se enumeran en la Tabla\(\PageIndex{1}\). Las células dendríticas son importantes centinelas que residen en la piel y membranas mucosas, que son portales de entrada para muchos patógenos. Los monocitos, macrófagos y células dendríticas son todos altamente fagocíticos e importantes promotores de la respuesta inmune a través de su producción y liberación de citocinas. Estas células proporcionan un puente esencial entre las respuestas inmunes innatas y adaptativas, como se discute en la siguiente sección así como en el siguiente capítulo.
| Tejido | Macrófagos |
|---|---|
| Cerebro y sistema nervioso central | Células microgliales |
| Hígado | Células Kupffer |
| Pulmones | Macrófagos alveolares (células de polvo) |
| Cavidad peritoneal | Macrófagos peritoneales |
Ejercicio\(\PageIndex{3}\)
- Describir las señales que activan las células asesinas naturales.
- ¿Cuál es la diferencia entre monocitos y macrófagos?
Conceptos clave y resumen
- Los elementos formados de la sangre incluyen glóbulos rojos (eritrocitos), glóbulos blancos (leucocitos) y plaquetas (trombocitos). De estos, los leucocitos están principalmente involucrados en la respuesta inmune.
- Todos los elementos formados se originan en la médula ósea como células madre (CMH) que se diferencian a través de la hematopoyesis.
- Los granulocitos son leucocitos caracterizados por un núcleo lobulado y gránulos en el citoplasma. Estos incluyen neutrófilos (PMN), eosinófilos y basófilos.
- Los neutrófilos son los leucocitos que se encuentran en mayor número en el torrente sanguíneo y principalmente combaten las infecciones bacterianas.
- Los eosinófilos se dirigen a las infecciones parasitarias. Los eosinófilos y basófilos están involucrados en las reacciones alérgicas. Ambos liberan histamina y otros compuestos proinflamatorios de sus gránulos tras la estimulación.
- Los mastocitos funcionan de manera similar a los basófilos pero se pueden encontrar en tejidos fuera del torrente sanguíneo.
- Las células asesinas naturales (NK) son linfocitos que reconocen y matan células anormales o infectadas al liberar proteínas que desencadenan la apoptosis.
- Los monocitos son leucocitos mononucleares grandes que circulan en el torrente sanguíneo. Pueden abandonar el torrente sanguíneo y tomar residencia en los tejidos corporales, donde se diferencian y se convierten en macrófagos específicos de tejido y células dendríticas.