
10.3: Estructura y Función del ARN
- Page ID
- 54501

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Objetivos de aprendizaje
- Describir la estructura bioquímica de los ribonucleótidos
- Describir las similitudes y diferencias entre ARN y ADN
- Describir las funciones de los tres tipos principales de ARN utilizados en la síntesis de proteínas
- Explicar cómo el ARN puede servir como información hereditaria
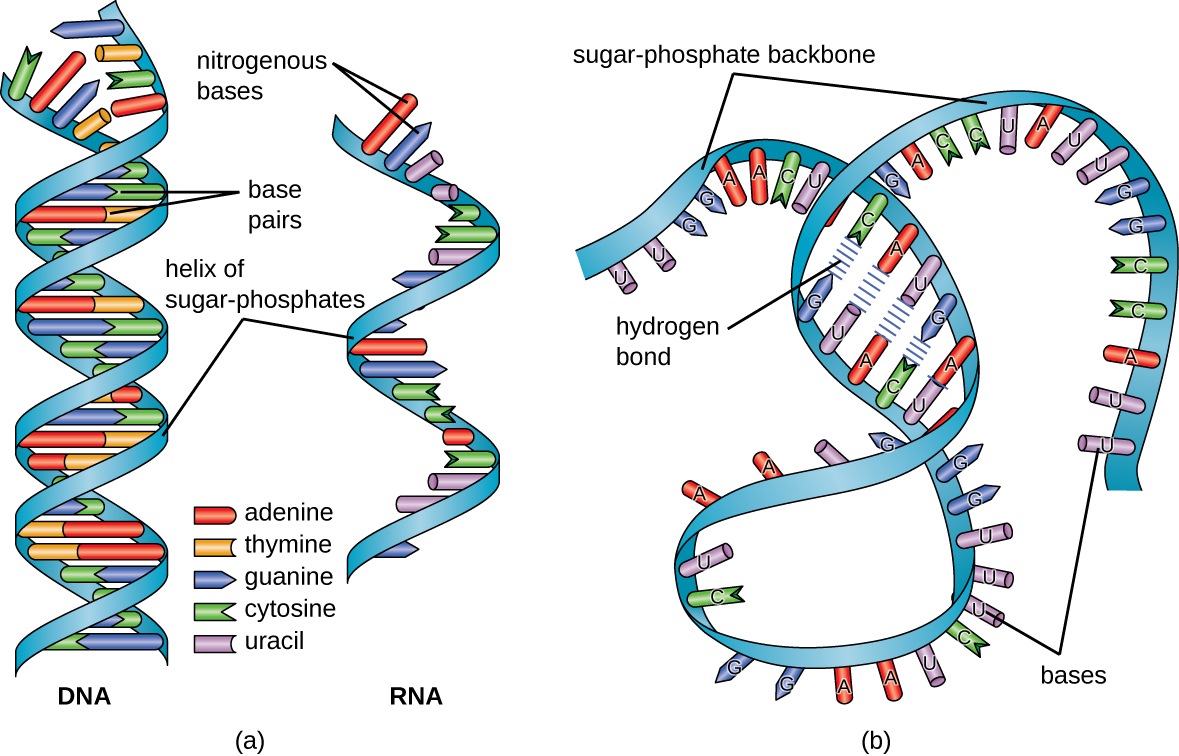
Estructuralmente hablando, el ácido ribonucleico (ARN), es bastante similar al ADN. Sin embargo, mientras que las moléculas de ADN son típicamente largas y bicatenarias, las moléculas de ARN son mucho más cortas y son típicamente monocatenarias. Las moléculas de ARN desempeñan una variedad de roles en la célula pero están involucradas principalmente en el proceso de síntesis de proteínas (traducción) y su regulación.
Estructura del ARN
El ARN es típicamente monocatenario y está hecho de ribonucleótidos que están unidos por enlaces fosfodiéster. Un ribonucleótido en la cadena de ARN contiene ribosa (el azúcar pentosa), una de las cuatro bases nitrogenadas (A, U, G y C) y un grupo fosfato. La sutil diferencia estructural entre los azúcares le da estabilidad agregada al ADN, lo que hace que el ADN sea más adecuado para el almacenamiento de información genética, mientras que la inestabilidad relativa del ARN lo hace más adecuado para sus funciones de más corto plazo.
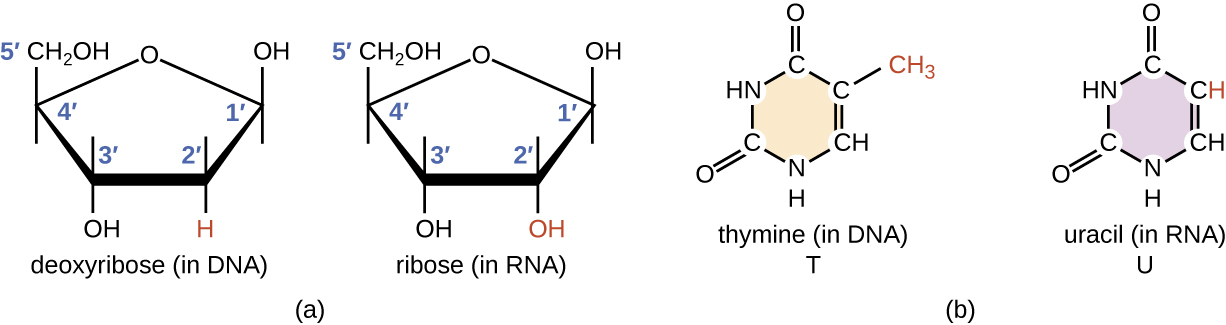
El uracilo de pirimidina específica de ARN forma un par de bases complementario con adenina y se usa en lugar de la timina utilizada en el ADN. Aunque el ARN es monocatenario, la mayoría de los tipos de moléculas de ARN muestran un amplio apareamiento de bases intramoleculares entre secuencias complementarias dentro de la cadena de ARN, creando una estructura tridimensional predecible esencial para su función (Figura\(\PageIndex{1}\) y Figura\(\PageIndex{2}\)).
Ejercicio\(\PageIndex{1}\)
¿En qué se diferencia la estructura del ARN de la estructura del ADN?
Funciones del ARN en la síntesis de proteínas
Las células acceden a la información almacenada en el ADN mediante la creación de ARN para dirigir la síntesis de proteínas a través del proceso de traducción. Las proteínas dentro de una célula tienen muchas funciones, incluyendo construir estructuras celulares y servir como catalizadores enzimáticos para reacciones químicas celulares que dan a las células sus características específicas. Los tres tipos principales de ARN directamente involucrados en la síntesis de proteínas son el ARN mensajero (ARNm), el ARN ribosómico (ARNr) y el ARN de transferencia (ARNt).
En 1961, los científicos franceses François Jacob y Jacques Monod plantearon la hipótesis de la existencia de un intermediario entre el ADN y sus productos proteicos, al que llamaron ARN mensajero. 1 La evidencia que respalda su hipótesis se recopiló poco después mostrando que la información del ADN se transmite al ribosoma para la síntesis de proteínas usando ARNm. Si el ADN sirve como la biblioteca completa de información celular, el ARNm sirve como fotocopia de la información específica necesaria en un momento determinado que sirve como instrucciones para elaborar una proteína.
El ARNm lleva el mensaje del ADN, que controla todas las actividades celulares en una célula. Si una célula requiere una determinada proteína para ser sintetizada, el gen para este producto se “enciende” y el ARNm se sintetiza a través del proceso de transcripción (ver Transcripción de ARN). El ARNm luego interactúa con ribosomas y otra maquinaria celular (Figura\(\PageIndex{3}\)) para dirigir la síntesis de la proteína que codifica durante el proceso de traducción (ver Síntesis de Proteínas). El ARNm es relativamente inestable y de corta duración en la célula, especialmente en células procariotas, asegurando que las proteínas solo se elaboran cuando es necesario.
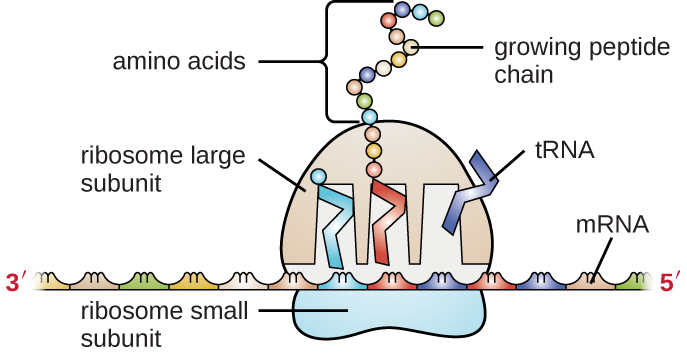
ARNr y ARNt son tipos estables de ARN. En procariotas y eucariotas, el ARNt y el ARNr se codifican en el ADN, luego se copian en moléculas de ARN largas que se cortan para liberar fragmentos más pequeños que contienen las especies de ARN maduras individuales. En eucariotas, la síntesis, corte y ensamblaje del ARNr en ribosomas se lleva a cabo en la región nucleolar del núcleo, pero estas actividades ocurren en el citoplasma de los procariotas. Ninguno de estos tipos de ARN lleva instrucciones para dirigir la síntesis de un polipéptido, pero juegan otros papeles importantes en la síntesis de proteínas.
Los ribosomas están compuestos por ARNr y proteína. Como su nombre indica, el ARNr es un constituyente principal de los ribosomas, que compone hasta aproximadamente el 60% del ribosoma en masa y proporciona la ubicación donde se une el ARNm. El ARNr asegura la alineación adecuada del ARNm, ARNt y los ribosomas; el ARNr del ribosoma también tiene una actividad enzimática (peptidil transferasa) y cataliza la formación de los enlaces peptídicos entre dos aminoácidos alineados durante la síntesis de proteínas. Aunque desde hace mucho tiempo se pensaba que el ARNr cumplía principalmente un papel estructural, su papel catalítico dentro del ribosoma se comprobó en el año 2000. 2 Científicos de los laboratorios de Thomas Steitz (1940—) y Peter Moore (1939—) de la Universidad de Yale pudieron cristalizar la estructura del ribosoma a partir de Haloarcula marismortui, un arqueón halófilo aislado del Mar Muerto. Debido a la importancia de este trabajo, Steitz compartió el Premio Nobel de Química 2009 con otros científicos que hicieron contribuciones significativas a la comprensión de la estructura del ribosoma.
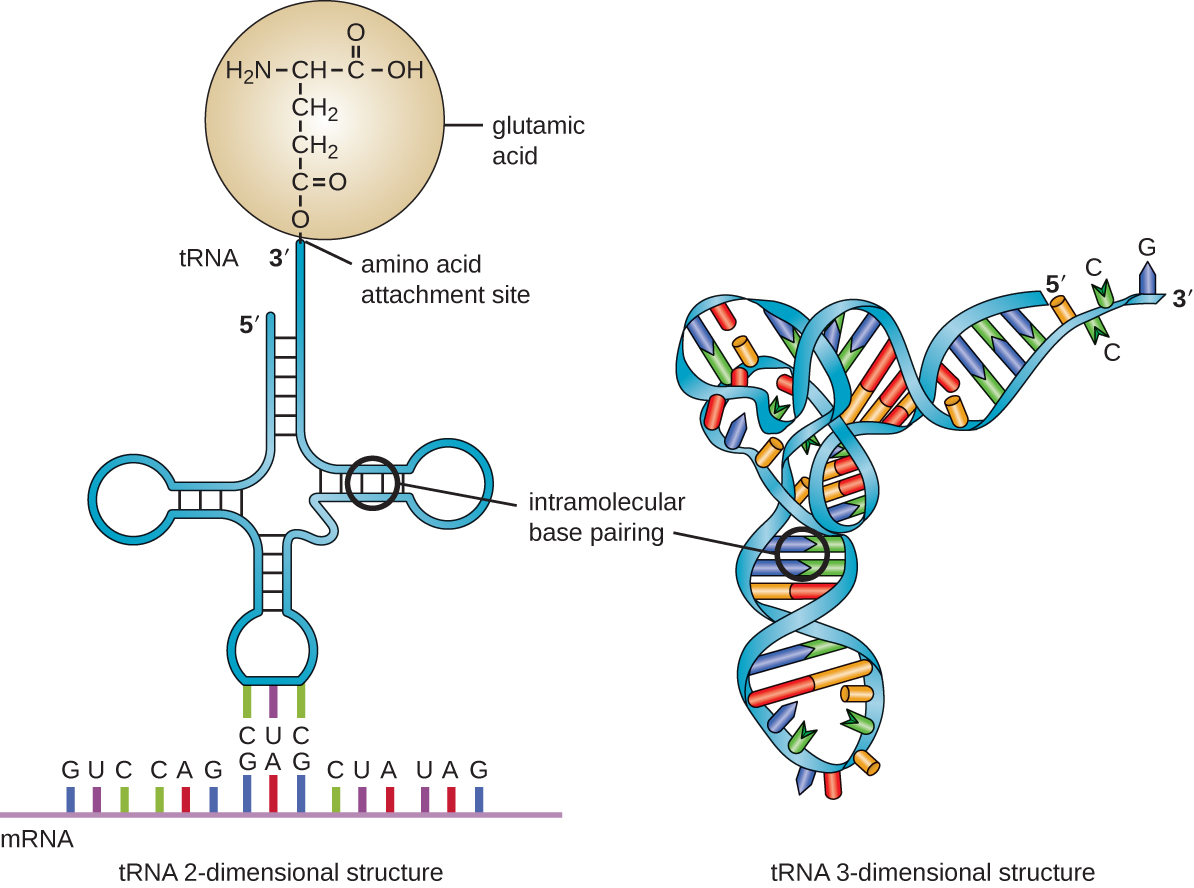
El ARN de transferencia es el tercer tipo principal de ARN y uno de los más pequeños, generalmente de solo 70—90 nucleótidos de longitud. Lleva el aminoácido correcto al sitio de síntesis de proteínas en el ribosoma. Es el emparejamiento de bases entre el ARNt y el ARNm lo que permite insertar el aminoácido correcto en la cadena polipeptídica que se está sintetizando (Figura\(\PageIndex{4}\)). Cualquier mutación en el ARNt o ARNr puede resultar en problemas globales para la célula debido a que ambos son necesarios para la síntesis de proteínas adecuada (Tabla\(\PageIndex{1}\)).
| mRNA | ARNr | ARNt | |
|---|---|---|---|
| Estructura | ARN monocatenario corto, inestable, correspondiente a un gen codificado dentro del ADN | Moléculas de ARN más largas y estables que componen el 60% de la masa del ribosoma | ARN corto (70-90 nucleótidos), estable con un extenso emparejamiento de bases intramoleculares; contiene un sitio de unión a aminoácidos y un sitio de unión a ARNm |
| Función | Sirve como intermediario entre el ADN y la proteína; utilizado por el ribosoma para dirigir la síntesis de la proteína que codifica | Asegura la alineación adecuada de ARNm, ARNt y ribosoma durante la síntesis de proteínas; cataliza la formación de enlaces peptídicos entre aminoácidos | Lleva el aminoácido correcto al sitio de síntesis de proteínas en el ribosoma |
Ejercicio\(\PageIndex{1}\)
¿Cuáles son las funciones de los tres tipos principales de moléculas de ARN involucradas en la síntesis de proteínas?
El ARN como información hereditaria
Aunque el ARN no sirve como la información hereditaria en la mayoría de las células, el ARN sí mantiene esta función para muchos virus que no contienen ADN. Por lo tanto, el ARN claramente tiene la capacidad adicional de servir como información genética. Aunque el ARN es típicamente monocatenario dentro de las células, existe una diversidad significativa en los virus. Los rinovirus, que causan el resfriado común; los virus de la influenza y el virus del Ébola son virus de ARN monocatenario. Los rotavirus, que causan gastroenteritis severa en niños y otros individuos inmunodeprimidos, son ejemplos de virus de ARN bicatenario. Debido a que el ARN bicatenario es poco común en las células eucariotas, su presencia sirve como indicador de infección viral. Las implicaciones para un virus que tiene un genoma de ARN en lugar de un genoma de ADN se discuten con más detalle en Virus.
Conceptos clave y resumen
- El ácido ribonucleico (ARN) es típicamente monocatenario y contiene ribosa como su azúcar pentosa y la pirimidina uracilo en lugar de timina. Una cadena de ARN puede experimentar un apareamiento de bases intramoleculares significativo para tomar una estructura tridimensional.
- Hay tres tipos principales de ARN, todos involucrados en la síntesis de proteínas.
- El ARN mensajero (ARNm) sirve como intermediario entre el ADN y la síntesis de productos proteicos durante la traducción.
- El ARN ribosómico (ARNr) es un tipo de ARN estable que es un constituyente principal de los ribosomas. Asegura la alineación adecuada del ARNm y los ribosomas durante la síntesis de proteínas y cataliza la formación de los enlaces peptídicos entre dos aminoácidos alineados durante la síntesis de proteínas.
- El ARN de transferencia (ARNt) es un tipo pequeño de ARN estable que transporta un aminoácido al sitio correspondiente de síntesis de proteínas en el ribosoma. Es el emparejamiento de bases entre el ARNt y el ARNm lo que permite insertar el aminoácido correcto en la cadena polipeptídica que se está sintetizando.
- Aunque el ARN no se utiliza para la información genética a largo plazo en las células, muchos virus sí utilizan el ARN como material genético.
Notas al pie
- 1 A. Rico. “La Era del Despertar del ARN: Biología Estructural del ARN en los Primeros Años”. Revisiones Trimestrales de Biofísica 42 núm. 2 (2009) :117—137.
- 2 P. Nissen et al. “La base estructural de la actividad ribosómica en la síntesis de enlaces peptídicos”. Ciencia 289 núm. 5481 (2000) :920—930.