Figura\(\PageIndex{1}\): Teodosio Dobzhansky, un importante científico que formuló la “síntesis moderna” del siglo XX reconciliando la teoría de la evolución de Charles Darwin y las ideas de Gregor Mendel sobre la herencia.
Después de 1950, reemplazando el concepto de “raza” como unidad de diversidad fue la “población”. Esto fue delineado por aquellos pioneros en la “nueva antropología física”, como Sherwood Washburn, Theodosius Dobzhansky y Julian Huxley, quienes tomaron prestada esta forma de enmarcar a grupos humanos de genetistas poblacionales contemporáneos (Figura 13.10). Las “razas” se definieron entonces simplemente como poblaciones que difieren en la frecuencia de algún gen o genes. Y, por otro lado, una “población” es un grupo de individuos potencialmente capaces de o realmente cruzarse debido a la proximidad geográfica, idioma, etnia, cultura y/o valores compartidos. Dicho de otra manera, una población es un grupo de mestizaje local con flujo génico reducido entre ellos y otros grupos de humanos. Se puede esperar que los miembros de una misma población compartan muchos rasgos genéticos (y, como resultado, muchos rasgos fenotípicos que pueden o no ser visibles externamente).
Figura\(\PageIndex{2}\): Julian Huxley (1942).
Pensar en los humanos en términos de poblaciones formaba parte de la “Síntesis Moderna” de Julian Huxley (1942), llamada así porque ayudó a conciliar dos principios fundamentales sobre la evolución que antes no habían tenido sentido juntos (Figura 13.11). Como se discutió en el capítulo 3, Gregor Mendel (1822-1884) pudo demostrar que la herencia estaba mediada por partículas discretas (o genes) y no mezclada en la descendencia. Sin embargo, fue difícil para algunos científicos del siglo XIX aceptar este modelo de herencia genética en ese momento porque gran parte de la variación biológica parecía ser continua y no particulada (tomar como ejemplos el color o la altura de la piel). En la década de 1930, se demostró que los rasgos podían ser poligénicos y que múltiples alelos podían ser responsables de cualquier rasgo fenotípico, produciendo así la variación continua en rasgos como el color de ojos que vemos hoy en día. Así, la “Síntesis Moderna” de Huxley describe no sólo cómo las poblaciones humanas son capaces de intercambiar genes a nivel microevolutivo sino también cómo múltiples alelos por un rasgo (intercambios poligénicos) pueden causar cambios macroevolutivos graduales.
La variación humana es clínica/continua (no discreta)
La diversidad humana no puede dividirse en “razas” discretas, porque la mayoría de los rasgos físicos varían de manera continua o “clinal”. Un ejemplo obvio de esto es cómo la altura humana no sólo viene en tres valores (“corto”, “medio” y “alto”) sino que varía a través de un espectro de alturas verticales alcanzables por humanos en todo el mundo. (Sin embargo, esto es con la única diferencia siendo la enorme divergencia en cómo factores como el tamaño corporal y rasgos como el color de la piel han sido vistos y utilizados sociopolíticamente como una forma de separar a las personas a lo largo de la historia). La necesidad de pasar de categorías tipológicas de “raza” a una comprensión más matizada de las poblaciones continuamente variables fue realizada por antropólogos que trabajaban en las décadas de 1960 y 1970, quienes cambiaron su enfoque hacia el estudio de los rasgos individuales en lugar del estudio de grupos (poblaciones, razas). Las evaluaciones sistemáticas de la variación biológica global en humanos solo comenzaron entonces, cuando se muestrearon grandes cantidades de loci genéticos para un gran número de muestras de poblaciones humanas distribuidas en todo el mundo. Fue durante la década de 1960 cuando se identificaron por primera vez los “clines” en la variación genética humana.
Frank B. Livingstone (1928-2005) escribió: “No hay razas, solo clines” (1962). Un clino es una gradación en la frecuencia de un alelo/rasgo entre poblaciones que viven en diferentes regiones geográficas. Para estudiar los rasgos humanos que están distribuidos clinalmente, a menudo se requiere realizar pruebas genéticas para descubrir las frecuencias verdaderas de un alelo o rasgo a través de un cierto espacio geográfico. Un ejemplo fácilmente visible de una distribución clinal vista en todo el mundo es el patrón de la variación humana en el color de la piel. Ya sea en el sur de Asia, África subsahariana o Australia, se encuentra piel marrón oscura. Los tonos de piel más pálidos se encuentran en poblaciones de latitudes más altas, como las que han vivido en áreas como Europa, Siberia y Alaska durante milenios. El color de la piel es fácilmente observable como un rasgo fenotípico que exhibe variación continua.
Una distribución clinal aún deriva de la herencia genética, pero los clinos a menudo corresponden a algún factor ambiental que cambia gradualmente. Los patrones clinales surgen cuando las presiones selectivas en un área geográfica difieren de las de otra, así como cuando las personas procrean y transmiten genes junto con sus vecinos más inmediatos. Existen varios mecanismos, selectivos y neutros, que pueden conducir a la distribución clinal de un alelo o un rasgo biológico. La selección natural es el mecanismo que produjo una clina global del color de la piel, por lo que el color de piel más oscuro protege a las poblaciones ecuatoriales de altas cantidades de radiación UV; hay una transición de disminución de la pigmentación en individuos que residen cada vez más lejos de los trópicos (Jablonski 2004; Jablonski y Chaplin 2000) (Figura 13.12). La capacidad e incapacidad para digerir la lactosa (azúcar de la leche) entre las diferentes comunidades del mundo varía según prácticas diferenciales e historias de consumo de leche y productos lácteos (Gerbault et al. 2011; Ingram et al. 2009). Donde la malaria parece ser más prevalente como estresante de enfermedad en poblaciones humanas, un gradiente clinal de creciente experiencia de anemia falciforme hacia estas regiones ha sido ampliamente estudiado por antropólogos genéticos (Luzzatto 2012). A veces, la selección de pareja culturalmente definida basada en algún rasgo observable también puede conducir a una variación clinal entre poblaciones.
Figura\(\PageIndex{3}\): Mapa global de colores de piel indígenas.
Dos procesos microevolutivos neutros que pueden producir una clina en un alelo o rasgo humano son el flujo génico y la deriva genética. Las formas en que los procesos neutros pueden producir distribuciones clinales se ve claramente al observar mapas clinales para diferentes grupos sanguíneos en el sistema de grupos sanguíneos ABO humanos (Figura 13.13). Por ejemplo, los científicos han identificado una clina de este a oeste en la distribución del alelo de tipo B de sangre a través de Eurasia. La frecuencia de portadores del alelo B disminuye gradualmente hacia el oeste cuando comparamos los grupos sanguíneos de las poblaciones del este y sudeste asiático con los de Europa. Esto muestra cómo las poblaciones que residen más cerca unas de otras tienen más probabilidades de cruzarse y compartir material genético (es decir, sufrir flujo génico). También vemos 90% ‒ 100% de individuos nativos sudamericanos, así como entre 70% ‒ 90% de los grupos aborígenes australianos, portando el alelo O (Mourant et al. 1976). Estas altas frecuencias probablemente se deben a la deriva genética aleatoria y a los efectos fundadores, en los que el tamaño de la población se redujo severamente por los primeros individuos portadores de alelo O que migraron a esas áreas. Con el tiempo, el tipo de sangre O se ha mantenido predominante.
Figura\(\PageIndex{4}\): Distribución global del grupo sanguíneo A.Figura\(\PageIndex{5}\): Distribución global del grupo sanguíneo B.Figura\(\PageIndex{6}\): Distribución global del tipo sanguíneo O.
La distribución de la variación humana: la diversidad genética es mayor dentro del grupo que entre grupos
Un problema con las clasificaciones basadas en la raza es que se basaban en una idea errónea de que las personas dentro de una categoría tipológica eran más similares entre sí que a las personas de otros grupos. En otras palabras, los conceptos de “raza” se basaron en la noción de que individuos con características particulares compartirían genes más similares entre sí dentro de una “raza” particular y compartirían menos con individuos de otras “razas” que poseen diferentes rasgos y maquillajes genéticos. Sin embargo, desde hace unos 50 años, estudios científicos han demostrado que la mayoría de las diferencias genéticas humanas a nivel mundial existen dentro de grupos (o “razas”) individualmente y no entre grupos.
Richard Lewontin (1929‒) es un biólogo y genetista evolutivo autor de un artículo que evalúa dónde radica la variación genética total en humanos. Este artículo, titulado “La distribución de la diversidad humana” (Lewontin 1972), abordó la siguiente pregunta: En promedio, ¿qué tan genéticamente similares son dos personas elegidas aleatoriamente del mismo grupo cuando se comparan con dos personas elegidas aleatoriamente de diferentes grupos? Lewontin estudió este problema mediante el uso de datos genéticos. Obtuvo datos para un gran número de diferentes poblaciones humanas a nivel mundial utilizando 17 marcadores genéticos (incluyendo alelos que codifican diversas enzimas y proteínas importantes, como las proteínas del grupo sanguíneo). El análisis estadístico que realizó utilizó una medida de las diferencias genéticas humanas en y entre poblaciones conocidas como índice de fijación (F ST). Técnicamente, F ST puede definirse como la proporción de varianza genética total dentro de una subpoblación en relación con la varianza genética total de una población entera. Por lo tanto, los valores F ST oscilan entre 0 y 1 (o, en ocasiones se verá esto expresado como un porcentaje entre 0% y 100%). Cuanto más cerca se acerca el valor F ST de una población (por ejemplo, la población mundial) a 1, mayor será el grado de diferenciación genética entre las subpoblaciones en relación con la población general. En su trabajo, Lewontin (1972) identificó que la mayoría de las diferencias genéticas humanas (85.4%) se encontraron dentro de subpoblaciones locales (e.g., los alemanes o isleños de Pascua), mientras que 8.3% se encontraron entre poblaciones dentro de grupos humanos continentales, y 6.3% fueron atribuibles a grupos tradicionales de “raza” (e.g.,” caucásico” o “amerindo”). Estos hallazgos han sido importantes para rechazar científicamente la existencia de razas biológicas (Long y Kittles 2008).
En 2002, otro artículo histórico de Noah Rosenberg y sus colegas (2002) exploró la variación genética humana en todo el mundo utilizando un conjunto de datos genéticos aún mayor. Utilizaron 377 marcadores altamente variables en el genoma humano y se muestrearon de 1,056 individuos representativos de 52 poblaciones. Los marcadores elegidos para el estudio no fueron los que codifican para ningún gen expresado. Debido a que estas regiones del genoma humano estaban hechas de genes no expresados, podemos entender estos marcadores como derivados neutralmente (a diferencia de derivados selectivamente) ya que no codifican ventajas o desventajas funcionales. Estos marcadores genéticos neutros probablemente reflejan una intrincada combinación de efectos fundadores regionales e historias poblacionales. Los análisis de estos marcadores neutros permitieron a los científicos identificar que la mayoría de la varianza genética global (93% ‒ 95%) se puede explicar por diferencias dentro de la población en los 377 loci genéticos, mientras que solo una pequeña proporción de varianza genética (3% ‒ 5%) puede atribuirse a diferencias entre grupos principales ( Rosenberg et al. 2002). Al igual que los hallazgos de Lewontin (1972), esto apoya la teoría de que no existen distintas razas biológicas, aunque los conceptos equivocados de raza puedan tener consecuencias sociales y políticas reales.
Los datos biológicos se ajustan a modelos de aislamiento por distancia y fuera de África
Otra nota es que si bien la población mundial puede dividirse genéticamente en “grupos”, “subconjuntos”, “grupos” o “racimos” que reflejan cierto grado de similitud genética, es más probable que estos clústeres identificables reflejen distancias genéticas o geográficas, ya sea con flujo génico facilitado por proximidad entre poblaciones o impedida por obstáculos como océanos o hábitats ambientalmente desafiantes (Rosenberg et al. 2005). En ocasiones, los grupos inferidos que utilizan múltiples loci genéticos son interpretados por los no genetistas literalmente como “poblaciones ancestrales”. No obstante, sería erróneo suponer a partir de estos resultados genéticos que alguna vez existieron grupos ancestrales altamente diferenciados y “puros”. Estas agrupaciones reflejan diferencias que han surgido a lo largo del tiempo debido a patrones clinales, deriva genética y/o flujo génico restringido o no restringido (Weiss y Long 2009). Los clústeres identificados por los científicos son arbitrarios y los parámetros utilizados para dividir la población global en grupos son subjetivos y dependen de las preguntas o distinciones particulares que se pongan en foco (Relethford 2009).
Adicionalmente, investigaciones sobre diversidad genética mundial han demostrado que la variación humana disminuye con el aumento de la distancia del África subsahariana, donde hay evidencia de que esta vasta región es el origen geográfico de humanos anatómicamente modernos (Liu et al. 2006; Prugnolle et al. 2005) (Figura 13.14). La diferenciación genética disminuye en los grupos humanos a medida que se muestrean los datos relativos al África subsahariana debido a los efectos de los fundadores en serie (Relethford 2004). A lo largo de la colonización humana del resto del mundo fuera de África, las poblaciones se separaron en olas en expansión a través de los continentes hacia Asia occidental, luego Europa y Asia oriental, seguidas de Oceanía y América. Como resultado, ocurrieron eventos fundadores en los que se perdió la variación genética, ya que la colonización de cada nueva región geográfica involucró a un número menor de individuos que se trasladaron de la población original más grande para establecer una nueva (Relethford 2004). La mayor variación genética se encuentra entre poblaciones que residen en diferentes partes del África subsahariana, mientras que otras poblaciones actuales en lugares como el norte de Europa y el extremo sur de América del Sur exhiben algunas de las menos diferenciación genética en relación con todas las poblaciones globales.
Además de encajar muy bien en el , la variación genética humana mundial se ajusta a un , que predice que la similitud genética entre los grupos disminuirá exponencialmente a medida que la distancia geográfica entre ellos aumenta. Esto se debe a las mayores y mayores restricciones al flujo de genes que presenta la distancia geográfica, así como a las diferencias culturales y lingüísticas que se dan como resultado de ciertos grados de aislamiento. Dado que los datos genéticos se ajustan a los modelos de aislamiento por distancia y fuera de África, estos hallazgos apoyan la abolición de agrupaciones “raciales”. Esta investigación demuestra que la variación humana es continua y no se puede diferenciar en categorías geográficamente discretas. No hay diferencias “inherentes” o “innatas” entre los grupos humanos; en cambio, la variación deriva de algún grado de selección natural, así como procesos neutros como el embotellamiento poblacional (Figura 13.15), mutaciones aleatorias en el ADN, deriva genética y flujo génico entre- mestizaje mate.
Figura\(\PageIndex{8}\): El efecto fundador es un cambio en el acervo genético de una pequeña población debido a un número limitado de individuos que se separan de una población parental.
Los humanos tienen mayor homogeneidad en comparación con muchas otras especies
Un dato importante a tener en cuenta es que los humanos son 99.9% idénticos entre sí. Esto significa que las distribuciones de la diversidad humana discutidas anteriormente solo conciernen a ese diminuto 0.1% de diferencia que existe entre todos los humanos a nivel mundial. En comparación con otras especies de mamíferos, incluidos los otros grandes simios, la diversidad humana es notablemente menor. Esto puede ser sorprendente dado que la población humana mundial ya ha superado los siete mil millones, y, al menos a nivel de superficie, parecemos ser bastante fenotípicamente diversos. Los enfoques moleculares de la genética humana y de primates nos dicen que las diferencias externas son meramente superficiales. Para una adecuada apreciación de la diversidad humana, tenemos que mirar a nuestros parientes más cercanos en el orden de los primates y la clase de mamíferos. En comparación con los chimpancés, gibones e incluso lobos grises y pandas gigantes, los humanos tienen una heterogeneidad promedio notablemente baja en todo el genoma.
Cuando observamos la diversidad genética de los chimpancés, es fascinante que los grupos de chimpancés occidentales, centrales, orientales y cameruneses tengan sustancialmente más diversidad genética entre ellos que grandes muestras globales de ADN humano (Bowden et al. 2012) (Figura 13.16). Esto es sorprendente dado que todos estos grupos de chimpancés viven relativamente cerca unos de otros en África, mientras que las mediciones de la diversidad genética humana se han realizado utilizando muestras de continentes completamente diferentes. Primero, los genetistas suponen que esto podría reflejar experiencias diferenciales del efecto fundador entre humanos y chimpancés. Debido a que todas las poblaciones humanas no africanas descendieron de un pequeño número de humanos anatómicamente modernos que abandonaron África, se esperaría que todos los grupos descendientes de ese grupo ancestral más pequeño fueran similares genéticamente. Segundo, nuestra especie es realmente joven, dado que solo hemos existido en el planeta por alrededor de 150,000 a 300,000 años. Esto dio a los humanos poco tiempo para que ocurrieran mutaciones genéticas aleatorias a medida que los genes se transmiten a través del mestizaje genético y la meiosis. Los chimpancés, sin embargo, han habitado diferentes nichos ecológicos, y se ha producido menos mestizaje entre los cuatro grupos de chimpancés en los últimos seis a ocho millones de años en comparación con la cantidad de flujo génico que se produjo entre las poblaciones humanas mundiales (Bowden et al. 2012).
Los avances recientes ahora han permitido el logro de muestras genéticas de la familia más grande de grandes simios y la evaluación de la diversidad genética entre bonobos, orangutanes y gorilas junto a la de chimpancés y humanos (Prado-Martinez et al. 2013). La recolección de estos datos y el análisis de la diversidad genética de los primates ha sido importante no solo para dilucidar cómo diferentes factores ecológicos, demográficos y climáticos han dado forma a nuestra evolución, sino también para informar sobre los esfuerzos de conservación y la investigación médica. Los genes que pueden codificar las susceptibilidades genéticas a enfermedades tropicales que afectan a múltiples primates se pueden estudiar a través de métodos genómicos. Las diferencias de especies en los genomas asociadas con el habla, el comportamiento o la cognición podrían decirnos más sobre cómo los individuos humanos pueden verse afectados por trastornos y afecciones neurológicas derivadas genéticamente o relacionadas con el habla (Prado-Martinez et al. 2013; Staes et al. 2017). En 2018, un estudio genómico de grandes simios también reportó diferencias genéticas entre chimpancés y humanos relacionadas con las divisiones celulares cerebrales (Kronenberg et al. 2018). A partir de estos resultados, se puede inferir que la variación cognitiva o conductual entre los humanos y los grandes simios podría relacionarse con un mayor número de neuronas corticales que se forman durante el desarrollo del cerebro humano (Kronenberg et al. 2018). Los estudios comparativos de la variación genética de los grandes simios humanos y no humanos destacan las complejas interacciones de las historias poblacionales, los cambios ambientales y la selección natural entre y dentro de las especies. Cuando se ve en el contexto de la diversidad general de los grandes simios, podemos reconsiderar cuán variable es relativamente la especie humana y cuán injustificados eran realmente los conceptos anteriores de “raza”.
Rasgos fenotípicos que reflejan la evolución neutra
La mayoría de los rasgos humanos son no concordantes. “” es un término que se usa para describir cómo los rasgos biológicos varían independientemente unos de otros, es decir, no se heredan de manera correlativa con otros rasgos genéticamente controlados. Por ejemplo, si supieras que un individuo tenía genes que codificaban para estatura alta, no podrías predecir si son de piel más clara o tienen el pelo rojo. Dependiendo del rasgo que se observe, se pueden encontrar diferentes patrones de variación fenotípica dentro y entre grupos en todo el mundo. En esta subsección se discutirán algunos rasgos fenotípicos que reflejan los patrones de variación genética mencionados anteriormente.
Figura\(\PageIndex{10}\): Cráneos humanos en Tana Toraja (Indonesia), escenario común en fosas públicas.
Mirando brevemente más allá de la variación genética, estudios recientes han revisitado temas anteriores de la antropología biológica de rasgos externamente observables, como la forma del cráneo (Figura 13.17). En los últimos 20 años, los antropólogos han evaluado el nivel al que la diversidad de formas craneales humanas refleja los resultados de los marcadores genéticos, como los utilizados previamente para adaptarse a modelos fuera de África (Relethford 2004) o los utilizados en la distribución de la diversidad humana entre y dentro de grupos (Lewontin 1972; Rosenberg et al. 2002). Usando tamaños de muestra más grandes de datos craneales recopilados de miles de cráneos en todo el mundo y una larga lista de mediciones craneales, los estudios demuestran una disminución similar en la diversidad con la distancia de África y muestran que la mayoría de la variación craneal ocurre dentro de poblaciones y no entre poblaciones ( Betti et al. 2009; Betti et al. 2010; Manica et al. 2007; Relethford 2001; von Cramon-Taubadel y Lycett 2008). La mayor diversidad craneal se encuentra entre cráneos de origen africano subsahariana, mientras que la menor variación se encuentra entre poblaciones que habitan lugares como Tierra del Fuego en el extremo sur de Argentina y Chile. Mientras que los pensadores antiguos e históricos pensaban anteriormente que las categorías de “raza” podrían determinarse razonablemente en función de las dimensiones del cráneo, los análisis modernos que utilizan conjuntos más informativos de rasgos craneales simplemente muestran que las migraciones fuera de África y las distancias relativas entre poblaciones pueden explicar la mayoría de diversidad craneal mundial (Betti et al. 2009).
Figura\(\PageIndex{11}\): Diagrama del laberinto óseo en el oído interno.
Este mismo patrón en variación fenotípica se ha encontrado incluso en estudios que examinan la variación de forma de la pelvis (Betti et al. 2013; Betti et al. 2014), los dientes (Rathmann et al. 2017) y el laberinto óseo humano de la oreja (Ponce de León et al. 2018) (Figura 13.18). La morfología esquelética de estos huesos todavía varía en todo el mundo, pero una mayor proporción de esa variación aún se puede atribuir a las formas en que las poblaciones humanas migraron por todo el mundo e intercambiaron genes con aquellos más cercanos a ellos en lugar de aquellos más alejados. La variación esquelética humana en estas partes del cuerpo es continua y no discreta. Dadas las importantes funciones del cráneo y estas otras partes esqueléticas, podemos inferir que los genes que sustentan su desarrollo han sido relativamente conservados por procesos evolutivos neutros como la deriva genética y el flujo génico. También es importante señalar que si bien algunos rasgos como la altura, el peso, las dimensiones craneales y la composición corporal están determinados, en parte, por genes, los procesos de desarrollo subyacentes detrás de estos rasgos están sustentados por mecanismos poligénicos complejos que han llevado al espectro continuo de variación en tales variables entre las poblaciones humanas de hoy en día.
Rasgos fenotípicos que reflejan la selección natural
A pesar de que 99.9% de nuestro ADN es el mismo entre todos los humanos en todo el mundo, y muchos rasgos reflejan procesos neutros, hay partes de ese 0.1% restante del genoma humano que codifican para diferencias individuales y regionales. De manera similar a los análisis craneométricos que se han realizado en las últimas décadas, la variación humana en el color de la piel también se ha reevaluado utilizando nuevos métodos y a la luz de un mayor conocimiento de la evolución biológica.
Las nuevas tecnologías permiten a los científicos utilizar fotometría de color para muestrear y cuantificar la longitud de onda visible del color de la piel, de una manera que los lectores de los siglos XIX y XX no pudieron. En un reporte, se encontró que 87.9% de la variación global del color de la piel puede atribuirse a diferencias genéticas entre grupos, 3.2% a las de las poblaciones locales dentro de las regiones y 8.9% dentro de las poblaciones locales (Relethford 2002). Esta distribución difiere significativamente y es la situación inversa que se encuentra en la distribución de las diferencias genéticas que vemos cuando examinamos marcadores genéticos como los alelos relacionados con el tipo de sangre. Sin embargo, este patrón de color de piel humana a nivel mundial no es sorprendente, dado que ahora entendemos que la selección pasada ha ocurrido para pieles más oscuras cerca del ecuador y pieles más claras en latitudes más altas (Jablonski 2004; Jablonski y Chaplin 2000). Si bien la mayoría de la diversidad genética refleja la variación neutra debido a las migraciones poblacionales, el aislamiento geográfico y la dinámica de flujo génico restringido, parte de la diversidad genética/fenotípica humana se explica mejor como adaptación local a las condiciones ambientales (es decir, selección). Dado que la variación del color de la piel es atípica en comparación con otros marcadores genéticos y rasgos biológicos, esto, de hecho, va en contra de tipologías de “raza” anteriores. Esto se debe a que estudios recientes muestran irónicamente cuán gran parte de la variación genética se relaciona con procesos neutros, mientras que el color de la piel no. De ello se deduce que el color de la piel no puede considerarse útil para hacer inferencias sobre otros rasgos humanos.
Además de las implicaciones sociales, la cuantificación e interpretación de la variación humana tiene importantes aplicaciones médicas y clínicas (National Research Council Committee on Human Genome Diversity 1997). Por ejemplo, estudios genómicos a gran escala de muestreo de poblaciones humanas distribuidas en todo el mundo han producido conocimiento detallado sobre la variación en la resistencia a enfermedades o susceptibilidad entre poblaciones y dentro de ellas Si pensamos en compañías farmacéuticas que desarrollan medicamentos para pacientes afroamericanos particularmente, la diversidad genética en predisposiciones a enfermedades y/o buena salud es probablemente mayor entre las personas de ascendencia africana de lo que estos negocios farmacéuticos han tenido en cuenta. A través del muestreo dirigido de varios grupos mundiales, los genetistas clínicos también pueden identificar factores de riesgo genéticos de ciertos trastornos comunes como enfermedades cardíacas crónicas, asma, diabetes, enfermedades autoinmunes y trastornos del comportamiento. Tener una comprensión de la biología específica de la población es crucial en el desarrollo de terapias, medicamentos y vacunas, ya que no todos los tratamientos pueden ser adecuados para cada ser humano, dependiendo de su genotipo. Durante el diagnóstico y tratamiento, es importante tener una perspectiva evolutiva sobre las relaciones gene-ambiente en los pacientes. Los conceptos tipológicos de “raza” no son útiles, dado que la mayoría de los grupos raciales (ya sean autoidentificados o no) popularmente reconocidos carecen de homogeneidad y son, de hecho, variables. La fibrosis quística, por ejemplo, ocurre en todas las poblaciones del mundo pero a menudo puede ser subdiagnosticada en poblaciones de ascendencia africana porque se considera una enfermedad “blanca” (Yudell et al. 2016).
Figura\(\PageIndex{12}\): El Laboratorio de Antropología Forense del Museo Nacional de Historia Natural, Instituto Smithsonian, Washington, D.C.
Por último, la asignación de “raza” a restos humanos es una práctica común en la antropología forense, especialmente en Estados Unidos y otros contextos mundiales donde se recuperan huesos y se asocian a investigaciones penales (Figura 13.19). Los antropólogos forenses han atribuido “raza” o ascendencia a conjuntos de esqueletos gracias a investigaciones científicas que han intentado dividir diferentes grupos humanos en categorías construidas culturalmente basadas en surtidos biológicamente “discretos” (Sauer 1992). En lugar de centrarse en las causas neutrales o selectivas de la variación biológica humana, la concentración en antropología forense se centra en lo probabilístico que puede ser asignar huesos de ciertas dimensiones a una de varias categorías raciales identificadas. Los antropólogos forenses no están de acuerdo con los conceptos tipológicos de “raza” del pasado sino que, en cambio, arraigan su categorización racial en métodos de estimación de probabilidad (Sauer 1992). A partir de muchas muestras de esqueletos de diferentes regiones del mundo, las pruebas estadísticas (como el análisis de función discriminante) les permiten distinguir la probabilidad de que ciertas dimensiones esqueléticas puedan predecir la ascendencia geográfica.
Es importante recordar que si bien es posible determinar el origen geográfico (o ascendencia) con base en la morfología del cráneo, nuevamente, la cantidad de distinción craneométrica requerida para distinguir si un cráneo pertenece a un grupo u otro tomará decisiones arbitrarias (Relethford 2009). Los individuos pueden variar en sus dimensiones esqueléticas según su origen continental, país de origen, origen regional, sexo, edad, factores ambientales y período de tiempo en el que vivieron, dificultando la asignación de individuos a categorías particulares de manera completamente significativa (Ousley et al. 2009). Cuando los informes forenses y artículos de revistas científicas dan una estimación de la ascendencia, es crucial tener en cuenta que las asignaciones responsables de ascendencia se realizarán a través de pruebas estadísticas sólidas y se declararán como una estimación de probabilidad. Hoy, también vivimos en un mundo más globalizado donde un individuo esquelético puede haber nacido originalmente de padres de dos categorías raciales tradicionales separadas. En contextos de gran heterogeneidad dentro de las poblaciones, esto definitivamente agrega dificultad al trabajo de científicos forenses y antropólogos preparando resultados para la sala de audiencias.


 Figura\(\PageIndex{1}\): Teodosio Dobzhansky, un importante científico que formuló la “síntesis moderna” del siglo XX reconciliando la teoría de la evolución de Charles Darwin y las ideas de Gregor Mendel sobre la herencia.
Figura\(\PageIndex{1}\): Teodosio Dobzhansky, un importante científico que formuló la “síntesis moderna” del siglo XX reconciliando la teoría de la evolución de Charles Darwin y las ideas de Gregor Mendel sobre la herencia. Figura\(\PageIndex{2}\): Julian Huxley (1942).
Figura\(\PageIndex{2}\): Julian Huxley (1942). Figura\(\PageIndex{3}\): Mapa global de colores de piel indígenas.
Figura\(\PageIndex{3}\): Mapa global de colores de piel indígenas.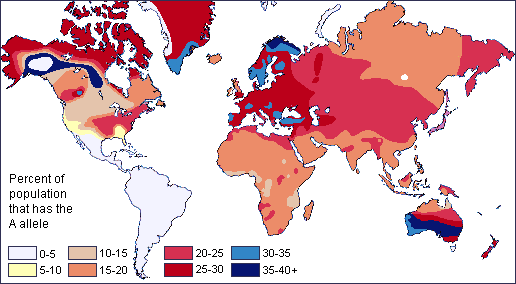.png) Figura\(\PageIndex{4}\): Distribución global del grupo sanguíneo A.
Figura\(\PageIndex{4}\): Distribución global del grupo sanguíneo A. Figura\(\PageIndex{5}\): Distribución global del grupo sanguíneo B.
Figura\(\PageIndex{5}\): Distribución global del grupo sanguíneo B. Figura\(\PageIndex{7}\): África subsahariana (sombreada oscuro/verde).
Figura\(\PageIndex{7}\): África subsahariana (sombreada oscuro/verde). Figura\(\PageIndex{8}\): El efecto fundador es un cambio en el acervo genético de una pequeña población debido a un número limitado de individuos que se separan de una población parental.
Figura\(\PageIndex{8}\): El efecto fundador es un cambio en el acervo genético de una pequeña población debido a un número limitado de individuos que se separan de una población parental. Figura\(\PageIndex{9}\): Chimpancé (Pan troglodytes).
Figura\(\PageIndex{9}\): Chimpancé (Pan troglodytes). Figura\(\PageIndex{10}\): Cráneos humanos en Tana Toraja (Indonesia), escenario común en fosas públicas.
Figura\(\PageIndex{10}\): Cráneos humanos en Tana Toraja (Indonesia), escenario común en fosas públicas. Figura\(\PageIndex{11}\): Diagrama del laberinto óseo en el oído interno.
Figura\(\PageIndex{11}\): Diagrama del laberinto óseo en el oído interno. Figura\(\PageIndex{12}\): El Laboratorio de Antropología Forense del Museo Nacional de Historia Natural, Instituto Smithsonian, Washington, D.C.
Figura\(\PageIndex{12}\): El Laboratorio de Antropología Forense del Museo Nacional de Historia Natural, Instituto Smithsonian, Washington, D.C.