
5.5: Modelos de población
- Page ID
- 70550

5.5. Modelos de población
Autores: A. Jan Hendriks y Nico van Straalen
Revisores: Aafke Schipper, John D. Stark y Thomas G. Preuss
Objetivos de aprendizaje
Deberías ser capaz de
- explicar los supuestos subyacentes a la modelización exponencial y logística de la población
- calcular la tasa intrínseca de crecimiento poblacional a partir de un conjunto dado de datos demográficos
- esbozar las conclusiones que pueden extraerse de la modelización poblacional en ecotoxicología
- indicar la posible contribución de los modelos poblacionales a la evaluación del riesgo químico
Palabras clave: tasa intrínseca de incremento, capacidad de carga, crecimiento exponencial,
Introducción
La evaluación del riesgo ecológico de los tóxicos generalmente se centra en los riesgos que corren los individuos, comparando las exposiciones con los niveles sin efecto. Sin embargo, en muchos casos no es la protección de plantas o animales individuales lo que es de interés sino la protección de una población viable de una especie en un contexto ecológico. La evaluación del riesgo generalmente no toma en cuenta la dinámica cuantitativa de las poblaciones y comunidades. Sin embargo, es urgente comprender y predecir los efectos de los productos químicos en niveles más allá del de los individuos por varias razones. Primero, necesitamos saber si los estándares de calidad son suficientemente pero no excesivamente protectores a nivel poblacional, cuando se extrapolan a partir de pruebas de toxicidad. En segundo lugar, las respuestas de cohortes aisladas y homogéneas en el laboratorio pueden ser diferentes de las de poblaciones heterogéneas interactuantes en el campo. Tercero, para establecer las prioridades correctas en la gestión, necesitamos conocer el efecto relativo y acumulativo de los químicos en relación con otras presiones ambientales.
Los modelos ecológicos de población de algas, macrófitos, invertebrados acuáticos, insectos, aves y mamíferos han sido ampliamente utilizados para abordar el riesgo de sustancias químicas potencialmente tóxicas, sin embargo, hasta hace poco, estos modelos solo se usaban raramente en el proceso de evaluación de riesgos regulatorios debido a la falta de conexión entre resultados del modelo y necesidades de evaluación de riesgos (Schmolke et al., 2010). Aquí, vamos a esbozar los principios básicos de la dinámica poblacional para aplicaciones de toxicología ambiental.
Crecimiento exponencial
Los libros de texto ecológicos suelen iniciar su capítulo sobre ecología poblacional introduciendo el crecimiento exponencial y logístico. Considerar una población de tamaño N. Si los recursos son ilimitados, y las tasas de nacimiento per cápita (b) y mortalidad (d) son constantes en una población cerrada a la migración, el número de individuos agregados a la población por unidad de tiempo (dN/dt) puede escribirse como:
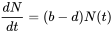 o
o 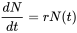
donde r se llama la tasa intrínseca de aumento. Esta ecuación diferencial se puede resolver con la condición de límite N (0) = N 0 para producir
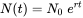
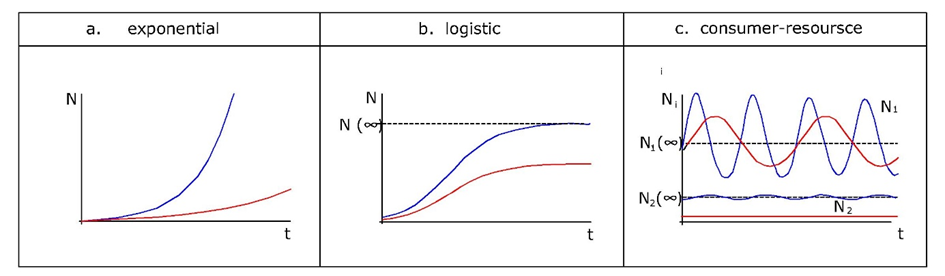
Dado que los tóxicos afectarán ya sea la reproducción o la supervivencia, o ambas, también afectarán la tasa de crecimiento exponencial (Figura 1a). Esto sugiere que r puede considerarse una medida del desempeño poblacional bajo estrés tóxico. Pero más que a partir de trayectorias poblacionales observadas, r suele estimarse a partir de datos de historia de vida. Sabemos por teoría demográfica básica que cualquier organismo con tasas vitales “invariables en el tiempo” (es decir, la fertilidad y la supervivencia pueden depender de la edad, pero no a tiempo), estará creciendo exponencialmente a la tasa r. La tasa intrínseca de aumento puede derivarse de las tasas de supervivencia y fertilidad específicas de la edad utilizando las llamadas Ecuación de Euler-Lotka, que dice:
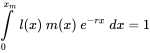
en el que x es edad, x m edad máxima, l (x) supervivencia desde la edad cero a la edad x y m (x) el número de crías producidas por unidad de tiempo a la edad x Desafortunadamente esta ecuación no permite una derivación simple de r; r debe obtenerse por iteración y el valor correcto es el que, al combinarse con los datos l (x) y m (x), hace que la integral sea igual a 1. Debido a esta complicación se suelen aplicar aproximaciones aproximadas. Por ejemplo, en muchos casos se puede obtener una estimación razonablemente buena para r a partir de la edad a la primera reproducción α, la supervivencia a la primera reproducción, S, y el rendimiento reproductivo, m, de acuerdo con la siguiente fórmula:
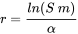
Esto se debe a que para muchos animales en el ambiente, especialmente aquellos con alto rendimiento reproductivo y baja supervivencia juvenil, la edad en la primera reproducción es la variable dominante que determina el crecimiento poblacional (Forbes y Calow, 1999).
El enfoque de modelización demográfica clásica, incluyendo la ecuación de Euler-Lotka, considera el tiempo como una variable continua y resuelve las ecuaciones por cálculo. Sin embargo, existe un formalismo equivalente basado en el tiempo discreto, en el que se supone que los eventos poblacionales ocurren sólo en momentos equidistantes. Las tasas vitales se resumen luego en una llamada matriz Leslie, una tabla de puntuaciones de supervivencia y fertilidad para cada clase de edad, organizada de tal manera que al multiplicarse por la distribución por edades en cualquier momento, se obtiene la distribución por edades en el siguiente punto temporal. Este tipo de modelado se presta más fácilmente a la simulación por computadora. El resultado es muy el mismo: si la matriz de Leslie es invariable en el tiempo la población crecerá cada vez paso por un factor λ, que se relaciona con r como ln λ = r (λ = 1 corresponde a r = 0). Matemáticamente hablando λ es el valor propio dominante de la matriz de Leslie. La ventaja de la versión de tiempo discreto es que λ se puede descomponer más fácilmente en sus partes componentes, es decir, los rasgos de historia de vida que se ven afectados por los tóxicos (Caswell, 1996).
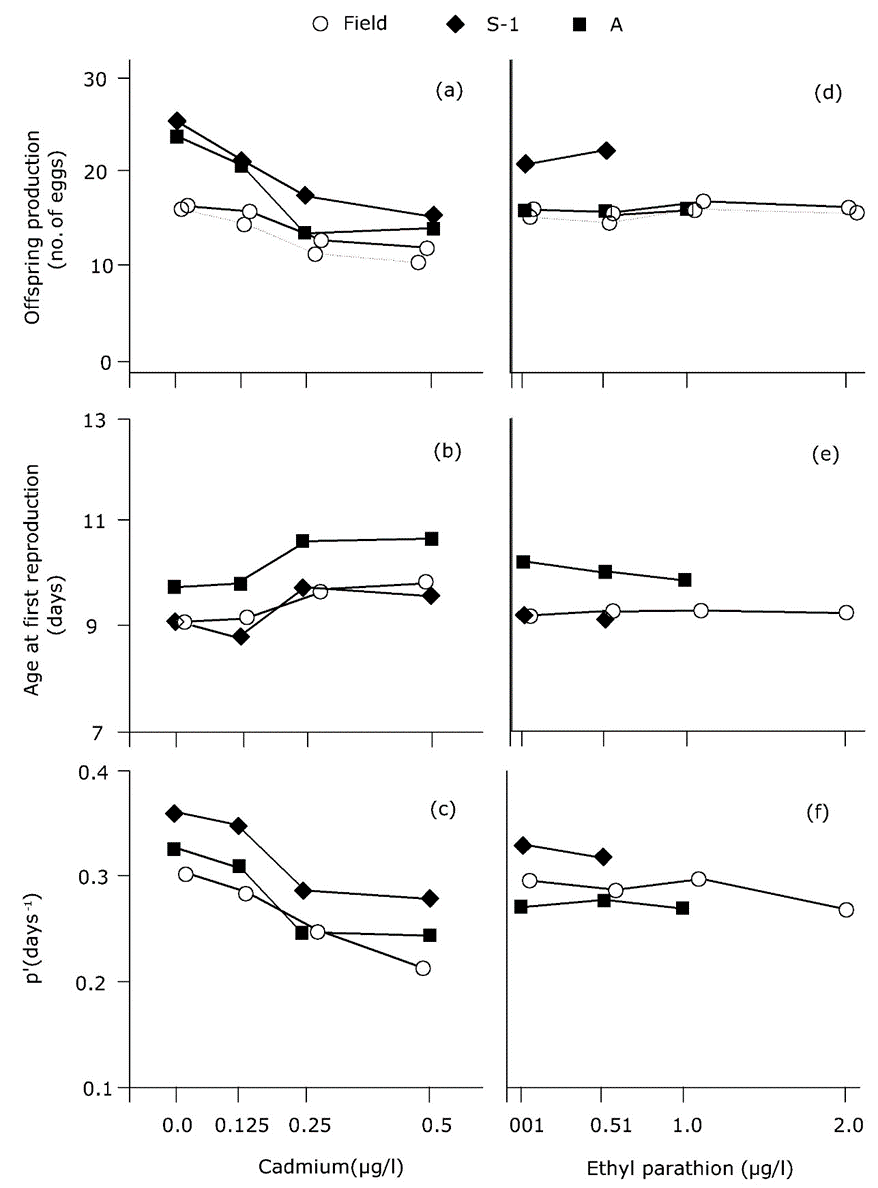
El enfoque demográfico para el crecimiento exponencial se ha aplicado numerosas veces en toxicología ambiental, con mayor frecuencia en estudios de pulgas de agua (Suhett et al., 2015) e insectos (Stark y Banks, 2003). Las pruebas se denominan “experimentos de respuesta de la tabla de vida” (ver sección Ecotoxicología poblacional en un entorno de laboratorio). El investigador observa los efectos de los tóxicos sobre la supervivencia y fertilidad específicas de la edad, y calcula r como medida del desempeño poblacional para cada concentración de exposición. Un ejemplo se da en la Figura 2, derivado de un estudio de Barata et al. (2000). Forbes y Calow (1999) concluyeron que el uso de r en ecotoxicología agrega relevancia ecológica al análisis, pero no necesariamente proporciona un punto final más sensible o menos sensible: r es tan sensible como las tasas vitales subyacentes a su estimación.
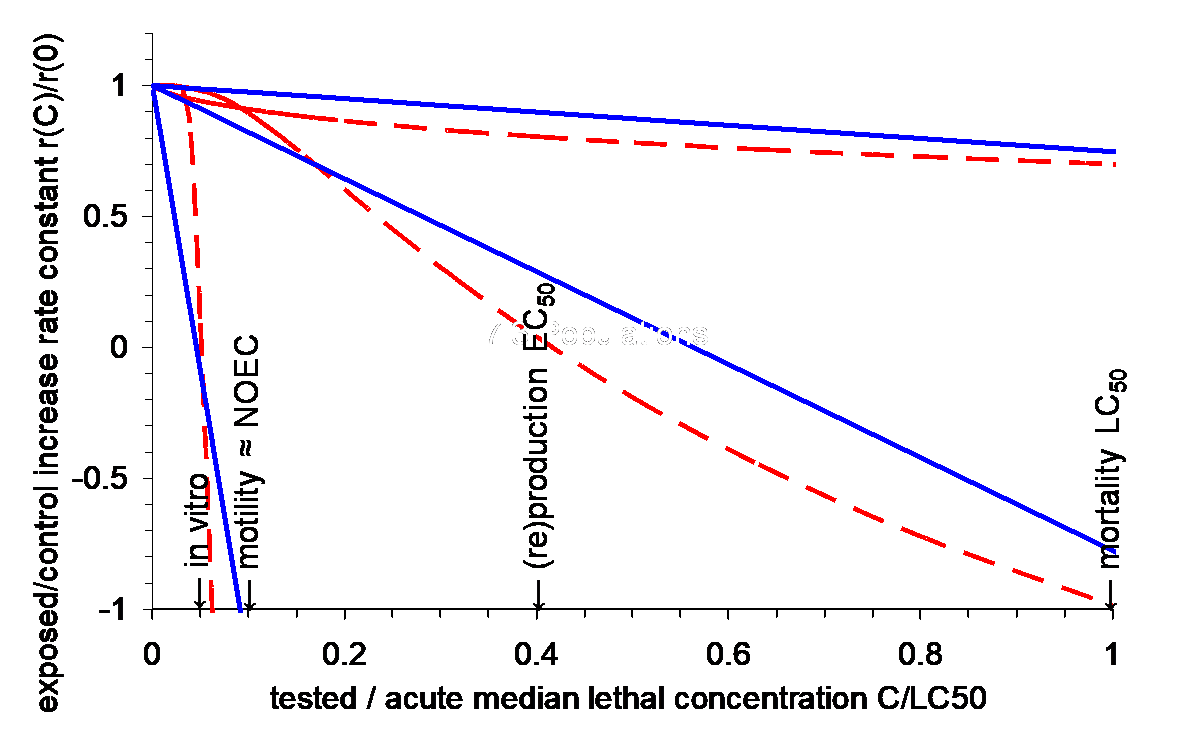
Hendriks et al. (2005) postularon que r debería mostrar una disminución casi lineal con la concentración de una sustancia química, escalada a la LC 50 (Figura 3). Esta relación se confirmó en un metaanálisis de 200 experimentos de laboratorio, en su mayoría relativos a especies de invertebrados (Figura 3). El apuntalamiento anecdótico para vertebrados grandes proviene de casos de campo donde la contaminación limita el desarrollo poblacional.
Crecimiento logístico
Como las poblaciones en crecimiento exponencial son obviamente raras, los modelos que incluyen alguna forma de dependencia de la densidad son más realistas. Un enfoque común es asumir que la tasa de natalidad b disminuye con la densidad debido a la creciente escasez de recursos. La suposición más simple es una disminución lineal con N, expresada de la siguiente manera:
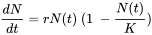
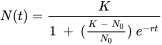
La pregunta es, ¿se pueden utilizar los parámetros de la ecuación de crecimiento logístico para medir el desempeño poblacional como en el caso del crecimiento exponencial? La aplicación práctica es limitada porque la capacidad de carga es difícil de medir en condiciones naturales y contaminadas. Muchas poblaciones de artrópodos en el campo, por ejemplo, fluctúan ampliamente debido a la dinámica depredador-presa, y casi nunca alcanzan su capacidad de carga dentro de una temporada de crecimiento. Un estudio experimental sobre la coleta elástica Folsomia candida (Noël et al., 2006) mostró que el zinc en la dieta no afectó la capacidad de carga de las poblaciones de laboratorio contenidas, aunque hubo varias interacciones por debajo de K que fueron influenciadas por el zinc, incluyendo hormesis (estimulación del crecimiento por dosis bajas de un tóxico), y efectos Allee (pérdida de potencial de crecimiento a baja densidad debido a menor tasa de encuentro).
Se espera que la dependencia de la densidad actúe como mecanismo de amortiguación a nivel poblacional porque la disminución de la población inducida por la toxicidad disminuye la competencia, sin embargo, los efectos dependen mucho de los detalles de la regulación poblacional. Esto se demostró en un modelo para halcón peregrino expuesto a DDE y PBDE (Schipper et al., 2013). Mientras que el tamaño de equilibrio de la población disminuyó por exposición tóxica, aumentó la probabilidad de que las aves individuales encontraran un territorio adecuado. Sin embargo, al mismo tiempo el número de aves no reproductoras que se desplazaban a la etapa de reproducción se volvió limitante y esto resultó en una fuerte disminución en el número de equilibrio de criadores.
Modelos de efectos mecanicistas
Para mejorar el potencial de aplicación de modelos poblacionales en la evaluación de riesgos, se deben incluir más detalles ecológicos de las especies bajo consideración, por ejemplo, efectos de dispersión, factores abióticos, depredadores y parásitos, dispersión, estructura del paisaje y muchos más. Un paso más es rastrear la fisiología y ecología de cada individuo en la población. Esto se hace en el enfoque de modelización dinámica del presupuesto energético (DEB) desarrollado por (Kooijman et al., 2009). Al incluir tales detalles, un modelo se volverá más realista y se podrán hacer predicciones más precisas sobre los efectos de las exposiciones tóxicas. Este tipo de modelos se denominan generalmente “modelos de efectos mecanicistas” (MEM). Permiten un vínculo causal entre la meta de protección, un escenario de exposición a tóxicos y los efectos adversos poblacionales generados por la producción del modelo (Hommen et al., 2015). La Autoridad Europea de Seguridad Alimentaria (EFSA) emitió en 2014 un documento de opinión que contiene directrices detalladas sobre el desarrollo de dichos modelos y cómo ajustarlos para que sean útiles en la evaluación de riesgos de los productos fitosanitarios.
Referencias
Caswell, H. (1996). La demografía cumple con la ecotoxicología: desenredando los efectos a nivel poblacional de sustancias tóxicas. En: Newman, M.C., Jagoe, C.H. (Eds.). Ecotoxicología. Un tratamiento jerárquico. Lewis Publishers, Boca Raton, pp. 255-292.
Barata, C., Baird, D.G., Amata, F., Soares, A.M.V.M. (2000). Comparación de la respuesta poblacional a contaminantes entre laboratorio y campo: una aproximación utilizando bancos de huevos ephippiales de Daphnia magna. Ecología Funcional 14, 513-523.
EFSA (2014). Opinión Científica sobre buenas prácticas de modelado en el contexto de modelos de efecto mecanicista para la evaluación de riesgos de productos fitosanitarios. Panel de Protección Vegetal y sus Residuos de la EFSA (PPR). Revista EFSA 12, 3589.
Forbes, V.E., Calow, P. (1999). Es la tasa de incremento per cápita una buena medida de los efectos a nivel poblacional en ecotoxicología. Toxicología y Química Ambiental 18, 1544-1556.
Hendriks, A.J., Maas, J.L., Heugens, E.H.W., Van Straalen, N.M. (2005). Metaanálisis de las tasas intrínsecas de incremento y capacidad de carga de poblaciones afectadas por factores tóxicos y otros factores estresantes. Toxicología y Química Ambiental 24, 2267-2277
Hommen, U., Forbes, V., Grimm, V., Preuss, T.G., Thorbek, P., Ducrot, V. (2015). Cómo utilizar modelos de efecto mecanicista en la evaluación de riesgos ambientales de plaguicidas: estudios de caso y recomendaciones del taller de SETAC Modelink. Evaluación y Gestión Ambiental Integrada 12, 21-31.
Kooijman, S.A.L.M., Baas, J., Bontje, D., Broerse, M., Van Gestel, C.A.M., Jager, T. (2009). Aplicaciones Ecotoxicológicas de la Teoría del Presupuesto Energético Dinámico. En: Devillers, J. (Ed.). Modelado Ecotoxicológico, Volumen 2, Springer, Dordrecht, pp. 237-260.
Noël, H.L., Hopkin, S.P., Hutchinson, T.H., Williams, T.D., Sibly, R.M. (2006). Hacia una ecología poblacional de ambientes estresados: los efectos del zinc en el coléculo Folsomia candida. Revista de Ecología Aplicada 43, 325-332.
Schipper, A.M., Hendriks, H.W.M., Kaufmann, M.J., Hendriks, A.J., Huijbregts, M.A.J. (2013). Modelación de interacciones de tóxicos y dependencia de la densidad en poblaciones de vida silvestre. Revista de Ecolog Aplicado y 50, 1469-1478.
Schmolke, A., Thorbek, P., Chapman, P., Grimm, V. (2010) Modelos ecológicos y evaluación del riesgo de plaguicidas: práctica actual de modelización. Toxicología y Química Ambiental 29, 1006-1012.
Stark, J.D., Banks, J.E. (2003) Efectos poblacionales de pesticidas y otros tóxicos sobre artrópodos. Revisión Anual de Entomología 48, 505-519.
Suhett, A.L. et al. (2015) Una visión general de la contribución de los estudios con cladóceros a la investigación del estrés ambiental. Acta Limnologica Brasiliensia 27, 145-159.