
3.5: Reglamento Inhibitorio de Competencia y Actividad
- Page ID
- 122879

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
La competencia inhibitoria juega un papel fundamental al permitirnos enfocarnos en algunas cosas a la vez, que luego podemos procesar de manera efectiva sin sobrecargarnos. La inhibición también asegura que aquellos detectores que sí se activan son los que están más excitados por una entrada dada; en términos evolutivos darwinianos, estos son los detectores más aptos.
Sin inhibición, la conectividad excitatoria bidireccional en la corteza causaría rápidamente que cada neurona se excitara mucho, porque no habría nada para verificar la propagación de la activación. Hay tantas conexiones excitatorias entre las neuronas que no tarda mucho en activarse cada neurona. Una buena analogía es colocar un micrófono cerca de un altavoz que está reproduciendo el sonido de ese micrófono — este es un sistema excitatorio bidireccional, y rápidamente lleva a ese chillido familiar y muy fuerte de “retroalimentación”. Si el sistema de audio de uno tuviera el equivalente del sistema inhibitorio en la corteza, en realidad sería capaz de evitar esta retroalimentación al bajar dinámicamente la ganancia de entrada en el micrófono, y/o el volumen de salida del altavoz.
Otra analogía útil es a un acondicionador de aire (AC), que tiene un control de termostato que determina cuándo entra en acción (y potencialmente qué tan fuerte es). Este tipo de sistema de control de retroalimentación permite que la habitación se caliente hasta un punto de ajuste determinado (por ejemplo, 75 grados F) antes de que comience a contrarrestar el calor. De manera similar, la inhibición en la corteza es proporcional a la cantidad de excitación, y produce un comportamiento de punto de ajuste similar, donde se evita que la actividad se vuelva demasiado alta: normalmente no más de aproximadamente el 15-25% de las neuronas en cualquier área dada están activas a la vez.
Sin embargo, la importancia de la inhibición va mucho más allá de esta función reguladora básica. La inhibición da lugar a la competencia: solo las neuronas más fuertemente excitadas son capaces de superar la señal de retroalimentación inhibitoria para activarse y enviar potenciales de acción a otras neuronas. Esta dinámica competitiva tiene numerosos beneficios en el procesamiento y el aprendizaje. Por ejemplo, la atención selectiva depende críticamente de la competencia inhibitoria. En el dominio visual, la atención selectiva es evidente cuando se busca un estímulo en una escena abarrotada (por ejemplo, la búsqueda de un amigo en una multitud como se describe en la introducción). No se puede procesar a todas las personas de la multitud a la vez, por lo que sólo unos relativamente pocos captan su atención, mientras que el resto son ignorados. En términos neuronales, decimos que los detectores para los pocos atendidos estaban lo suficientemente excitados como para superar a todos los demás, que permanecen por debajo del umbral de disparo debido a los altos niveles de inhibición. Tanto los factores de abajo hacia arriba como de arriba hacia abajo pueden contribuir a que los detectores neuronales superen el umbral o no, pero sin inhibición, no habría ninguna capacidad de seleccionar solo unos pocos para enfocarse en primer lugar. Curiosamente, las personas con síndrome de Balint, que tienen daño bilateral en la corteza parietal (que juega un papel crítico en la atención espacial de este tipo), muestran efectos atencionales reducidos y también suelen ser incapaces de procesar nada si una exhibición visual contiene más de un elemento (es decir, “simultanagnosia “— la incapacidad de reconocer objetos cuando hay múltiples simultáneamente presentes en una escena). Exploraremos estos fenómenos en el Capítulo Percepción.
Veremos en el Capítulo de Aprendizaje que la competencia inhibitoria facilita el aprendizaje al proporcionar esta presión de selección, por lo que solo se activan los detectores más excitados, que luego se refuerzan a través del proceso de aprendizaje para hacer que los detectores más activos estén aún mejor sintonizados para los insumos actuales, y con ello más probabilidades de responder a ellos de nuevo en el futuro. Este tipo de bucle de retroalimentación positiva sobre episodios de aprendizaje lleva al desarrollo de muy buenos detectores para el tipo de cosas que tienden a surgir en el entorno. Sin la competencia inhibitoria, un gran porcentaje de neuronas se entrenaría para cada insumo, y no habría especialización de detectores para categorías específicas en el ambiente. Toda neurona terminaría detectando débilmente todo, y así no lograría nada. Así, nuevamente vemos que la competencia y las limitaciones en realidad pueden ser sumamente beneficiosas.
Un término resumido para los tipos de patrones neuronales de actividad que se desarrollan en presencia de competencia inhibitoria son las representaciones dispersas distribuidas. Estas tienen relativamente pocas (15-25%) neuronas activas a la vez, y por lo tanto estas neuronas están más sintonizadas para las entradas actuales de lo que estarían de otra manera en una representación completamente distribuida con niveles mucho más altos de actividad general. Así, aunque técnicamente la inhibición no contribuye directamente a las funciones básicas de procesamiento de información como la categorización, debido a que la conectividad inhibitoria es estrictamente local dentro de un área cortical dada, la inhibición juega un papel indirecto crítico en la conformación de patrones de actividad neuronal en cada nivel.
Feedforward e inhibición de retroalimentación
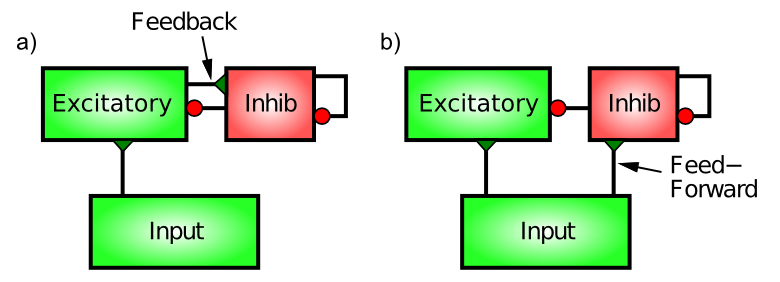
Hay dos patrones distintos de conectividad neural que impulsan interneuronas inhibitorias en la corteza, feedforward y feedback (Figura 3.16). Sólo para mantener las cosas interesantes, estas no son lo mismo que las conexiones entre las neuronas excitadoras. Funcionalmente, la inhibición de feedforward puede anticipar cuán excitadas se volverán las neuronas excitadoras, mientras que la retroalimentación refleja con precisión el nivel real de activación que logran.
La inhibición de retroalimentación es la más intuitiva, así que comenzaremos con ella. Aquí, las interneuronas inhibitorias son impulsadas por las mismas neuronas excitadoras a las que luego proyectan e inhiben. Este es el circuito clásico de “retroalimentación” del ejemplo de CA. Cuando un conjunto de neuronas excitadoras comienza a activarse, luego comunican esta activación a las interneuronas inhibitorias (a través de sinapsis glutamatérgicas excitadoras sobre interneuronas inhibitorias; las neuronas inhibidoras tienen que excitarse como todos los demás). Esta excitación de las neuronas inhibidoras hace que disparen potenciales de acción que regresan directamente a las neuronas excitadoras, abriendo sus canales iónicos inhibitorios a través de la liberación de GABA. La afluencia de iones Cl- (cloruro) de los canales de entrada inhibitorios en estas neuronas excitadoras actúa para hacerlas retroceder en la dirección del potencial impulsor inhibitorio (en la analogía del tira y afloja, el tipo inhibitorio se hace más grande y tira más fuerte). Así, la excitación engendra inhibición que contrarresta la excitación y mantiene todo bajo control, al igual que una ráfaga de aire frío de la unidad de CA.
La inhibición de Feedforward es quizás un poco más sutil. Opera cuando las entradas sinápticas excitatorias a las neuronas excitadoras en un área determinada también impulsan las interneuronas inhibitorias en esa área, provocando que las interneuronas inhiban a las neuronas excitatorias en proporción a la cantidad de entrada excitatoria que están recibiendo actualmente. Esto sería como un termostato reaccionando a la cantidad anticipada de calor, por ejemplo, encendiendo la CA en función de la temperatura exterior. Por lo tanto, la diferencia clave entre la inhibición de la alimentación directa y la retroalimentación es que la alimentación directa refleja la entrada excitatoria neta, mientras que la retroalimentación refleja la salida de activación real de un conjunto dado de neuronas excitadoras.
Como veremos en la exploración, la función anticipatoria de la inhibición de feedforward es crucial para limitar los tipos de oscilaciones dramáticas de retroalimentación que pueden desarrollarse en un sistema puramente impulsado por retroalimentación. Sin embargo, demasiada inhibición feedforward hace que el sistema sea muy lento para responder, por lo que hay un equilibrio óptimo de los dos tipos que da como resultado una dinámica inhibitoria muy robusta.
Exploración de la dinámica inhibitoria de las interneuronas
- Inhibición (inhib.proj): esta simulación muestra cómo las dinámicas inhibitorias de feedforward y feedback conducen al control robusto de las neuronas piramidales excitatorias, incluso en presencia de excitación bidireccional.
Función de inhibición de FFFB
Podemos implementar de manera eficiente la forma de inhibición feedforward (FF) y feedback (FB) sin requerir realmente las interneuronas inhibitorias, usando el ingreso neto promedio y los niveles de actividad en una capa dada, en una ecuación simple que se muestra a continuación. Esto funciona sorprendentemente bien, sin requerir la posterior adaptación de parámetros durante el aprendizaje, y esta forma de inhibición FFFB es ahora la predeterminada, reemplazando la forma de inhibición K-winners-Take-All (KwTA) utilizada en la 1ª Edición del libro de texto.
La entrada neta excitatoria promedio a una capa (o grupo de unidades dentro de una capa, si la inhibición está operando en ese nivel) es solo el promedio de la entrada neta (\(\eta_{i}\)) de cada unidad en la capa/grupo:
- \(<\eta>=\sum_{n} \frac{1}{n} \eta_{i}\)
Del mismo modo, la activación promedio es solo el promedio de los valores de activación (\(y_{i}\)):
- \(<y>=\sum_{n} \frac{1}{n} y_{i}\)
Calculamos la conductancia inhibitoria global aplicada uniformemente a todas las unidades de la capa/grupo con solo unos pocos parámetros clave aplicados a cada uno de estos dos promedios. Debido a que el componente de retroalimentación tiende a impulsar oscilaciones (alternativamente por encima y por debajo de la activación promedio), aplicamos una dinámica de integración de tiempo simple en ese término. El feedforward no requiere este tiempo de integración, pero sí requiere un término de compensación, el cual se determinó ajustando la inhibición real generada por nuestras ecuaciones anteriores de KwTA. Así, la conductancia inhibitoria global es solo la suma de los dos términos (ff y fb), con un factor de ganancia inhibitorio general gi:
- \(g_{i}(t)=\operatorname{gi}[\mathrm{ff}(t)+\mathrm{fb}(t)]\)
Este factor gi suele ser el único parámetro manipulado para determinar qué tan activa es en general una capa. Normalmente un valor de 1.5 es tan bajo como se usa, para dar un patrón de activación más ampliamente distribuido, siendo muy típicos valores alrededor de 2.0 (a menudo 2.1 o 2.2 funcionan mejor). Para capas muy dispersas (por ejemplo, una sola unidad de salida activa), se pueden usar valores de hasta alrededor de 3.5 o así.
El término feedforward (ff) es:
- \(\mathrm{ff}(t)=\mathrm{ff}[<\eta>-\mathrm{ff} 0]_{+}\)
donde ff es el factor de ganancia general para el componente feedforward (establecido en 1.0 por defecto), y ff0 es un desplazamiento (establecido en 0.1 por defecto) que se resta del valor de netinput promedio" src="https://grey.colorado.edu/mediawiki/...c90b756a54.png">.
El término feedback (fb) es:
- \(\mathrm{fb}(t)=\mathrm{fb}(t-1)+d t[\mathrm{fb}<y>-\mathrm{fb}(t-1)]\)
donde fb es el factor de ganancia general para el componente de retroalimentación (0.5 por defecto), dt es la constante de tiempo para integrar la inhibición de retroalimentación (0.7 por defecto), y el t-1 indica el valor anterior de la inhibición de retroalimentación; esta ecuación especifica un plegamiento gradual del nuevo factor de inhibición en la parte superior de lo que había antes, y el valor dt relativamente rápido de 0.7 hace que rastree el nuevo valor con bastante rapidez: hay un retraso suficiente para planchar las oscilaciones.
En general, debe quedar claro que esta inhibición de FFFB es extremadamente simple de calcular (mucho más simple que el cálculo anterior de KwTA), y se comporta de una manera mucho más proporcional en relación con el impulso excitatorio en las unidades —si hay mayor entrada excitatoria general, entonces el promedio la activación general en la capa será mayor, y viceversa. El cálculo anterior basado en Kwta tendió a ser más rígido e impuso un comportamiento más fuerte como un punto de ajuste. La dinámica FFFB, estando mucho más estrechamente ligada a la forma en que funcionan realmente las interneuronas inhibidoras, debería proporcionar una simulación biológicamente más precisa.
Exploración de la inhibición de FFFB
Para ver la inhibición de FFFB en acción, puede seguir las instrucciones de la última parte del modelo de inhibición (inhib.proj).