
4.2: Biología de la Plasticidad Sináptica
- Page ID
- 122872

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
El aprendizaje equivale a cambiar la eficacia sináptica general de la sinapsis que conecta dos neuronas. La sinapsis tiene muchas partes móviles (ver Neurón/Biología), cualquiera de las cuales podría ser potencialmente el factor crítico para hacer que su eficacia general cambie. ¿En cuántos se te ocurren? La búsqueda del factor (es) crítico (es) dominó la fase inicial de la investigación sobre plasticidad sináptica, y a lo largo de los años se han encontrado evidencias de la implicación de diversos factores, desde la cantidad de neurotransmisor presináptico liberado, hasta el número y eficacia de los receptores de AMPA postsinápticos, y cosas aún más sutiles como la alineación de componentes pre y postsinápticos, y cambios más dramáticos como la clonación de múltiples sinapsis. Sin embargo, el factor dominante para los cambios de aprendizaje duraderos parece ser el número y la eficacia de los receptores de AMPA postsinápticos.

La Figura 4.1 muestra los cinco pasos críticos en la cascada de eventos que impulsan el cambio en la eficacia del receptor AMPA. Los receptores NMDA y el ion calcio (Ca++) juegan un papel central: los canales de NMDA permiten que Ca++ ingrese a la columna postsináptica. En todas las células del cuerpo, el Ca++ suele desempeñar un papel importante en la regulación de la función celular, y en la neurona, es capaz de desencadenar una serie de reacciones químicas que terminan controlando cuántos receptores AMPA son funcionales en la sinapsis. Para obtener detalles sobre estas reacciones, consulte Biología Detallada del Aprendizaje. Esto es lo que se necesita para que el Ca++ entre en la celda postsináptica:
- El potencial de membrana postsináptica (Vm) debe ser elevado, como resultado de todas las entradas sinápticas excitatorias que ingresan a la célula. El contribuyente más importante a este nivel de Vm es en realidad el potencial de acción de retropropagación: cuando una neurona dispara un potencial de acción, no solo avanza hacia fuera del axón, sino también hacia atrás por las dendritas (a través de canales Na+ activos sensibles al voltaje a lo largo de las dendritas). Así, toda la neurona llega a conocer cuando se dispara —veremos que esto es increíblemente útil computacionalmente.
- La Vm elevada provoca que los iones magnesio (Mg+) sean repelidos (las cargas positivas se repelen entre sí) fuera de las aberturas de los canales NMDA, desbloqueándolos.
- La neurona presináptica dispara un potencial de acción, liberando neurotransmisor de glutamato en la hendidura sináptica.
- El glutamato se une al receptor NMDA, abriéndolo para permitir que los iones Ca++ fluyan hacia la célula postsináptica. Esto solo ocurre si el NMDA también está desbloqueado. Esta dependencia del NMDA tanto en la actividad pre como postsináptica fue una de las primeras pistas importantes sobre la naturaleza del aprendizaje, como vemos más adelante.
- La concentración de Ca++ en la columna postsináptica impulsa aquellas complejas reacciones químicas (Biología Detallada del Aprendizaje) que terminan cambiando el número y la eficacia de los receptores AMPA. Debido a que estos receptores AMPA proporcionan el impulso primario de entrada excitatoria en la neurona, cambiándolos cambia el efecto excitatorio neto de un potencial de acción presináptica sobre la neurona postsináptica. Esto es lo que se entiende por cambiar la eficacia sináptica, o peso.
El Ca++ también puede ingresar a la celda postsináptica a través de canales de calcio controlados por voltaje (VGCC) que son canales de calcio que solo se abren cuando el potencial de membrana está elevado. A diferencia de NMDA, sin embargo, no son sensibles a la actividad neural presináptica, solo dependen de la actividad postsináptica. Esto tiene importantes implicaciones computacionales, como discutiremos más adelante. Los VGCC contribuyen menos a los niveles de Ca++ que NMDA, por lo que NMDA sigue siendo el jugador dominante.
Los receptores metabotrópicos de glutamato (mGlu) también juegan un papel importante en la plasticidad sináptica. Estos receptores no permiten que los iones fluyan a través de la membrana (es decir, no son ionotrópicos), y en su lugar desencadenan directamente reacciones químicas cuando el neurotransmisor se une a ellos. Estas reacciones químicas pueden entonces modular los cambios en los receptores AMPA desencadenados por Ca++.
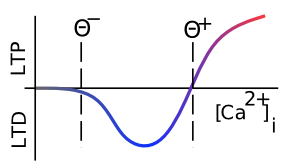
Hemos estado hablando de cambios en la eficacia del receptor AMPA sin especificar en qué dirección cambian. Potenciación a Largo Plazo (LTP) es el término biológico para aumentos duraderos en la eficacia de AMPA, y la Depresión a Largo Plazo (LTD) significa disminuciones duraderas en la eficacia de AMPA. Durante mucho tiempo, los investigadores se enfocaron principalmente en LTP (que generalmente es más fácil de inducir), pero finalmente se dieron cuenta de que ambas direcciones de plasticidad sináptica son igualmente importantes para el aprendizaje. La Figura 4.2 muestra cómo esta dirección de cambio depende del nivel general de Ca++ en la columna postsináptica (acumulado en unos pocos 100 de milisegundos al menos — las constantes de tiempo relevantes para los efectos del Ca++ sobre la plasticidad sináptica son bastante lentas) — los niveles bajos impulsan el LTD, mientras que los niveles altos producir LTP. Esta propiedad será crítica para nuestro modelo computacional. Tenga en cuenta que el retraso en los efectos de plasticidad sináptica basados en los niveles de Ca++ significa que la sinapsis no siempre tiene que hacer LTD en su camino hasta LTP; hay tiempo para que el Ca++ alcance un nivel alto para impulsar LTP antes de que los pesos comiencen a cambiar.
Aprendizaje hebbio y canales NMDA
El famoso psicólogo canadiense Donald O. Hebb predijo la naturaleza del canal NMDA muchos años antes de su descubrimiento, con solo pensar en cómo debería funcionar el aprendizaje a nivel funcional. Aquí hay una cita clave:
- Supongamos que la persistencia o repetición de una actividad reverberadora (o “rastro”) tiende a inducir cambios celulares duraderos que se suman a su estabilidad... Cuando un axón de la célula A está lo suficientemente cerca como para excitar una célula B y de manera repetida o persistente participa en su disparo, algún proceso de crecimiento o metabólico el cambio tiene lugar en una o ambas celdas de tal manera que se incrementa la eficiencia de A, ya que una de las celdas que dispara B.
Esto se puede resumir de manera más concisa como celdas que se disparan juntas, se unen por cable. El canal NMDA es esencial para este proceso, ya que requiere tanto actividad pre como postsináptica para permitir que Ca++ ingrese e impulse el aprendizaje. Puede detectar la coincidencia del disparo neural. Curiosamente, Hebb tiene fama de haber dicho algo en el sentido de “gran cosa, sabía que ya tenía que ser así” cuando alguien le dijo que su principio de aprendizaje había sido descubierto en la forma del receptor NMDA.
Matemáticamente, podemos resumir el aprendizaje hebbio como:
- \(\Delta w=x y\)
donde\(\Delta w\) es el cambio en el peso sináptico w, en función del envío de actividad\(x\) y recepción de actividad\(y\).
Cada vez que ves este tipo de producto pre-post en una regla de aprendizaje, tiende a describirse como una forma de aprendizaje hebbio. Para un tratamiento más detallado del aprendizaje hebbio y diversas variantes populares del mismo, consulte Hebbian Learning.
Como detallaremos a continuación, esta forma más básica de aprendizaje hebbio es muy limitada, porque los pesos solo subirán (dado que las actividades neuronales son tasas de picos y por lo tanto solo cantidades positivas), y lo harán sin ataduras. Curiosamente, el propio Hebb solo parecía haber contemplado LTP, no LTD, así que quizás esto sea adecuado. Pero no va a hacer nada útil en un modelo computacional. Antes de llegar al lado computacional de las cosas, cubrimos un resultado más importante en la biología.
Plasticidad dependiente de sincronización de picos

La Figura 4.3 muestra los resultados de un experimento realizado por Bi y Poo en 1998 que capturó la imaginación de muchos científicos, y ha resultado en un extenso trabajo de modelado computacional. Este experimento mostró que el orden preciso de disparo entre una neurona pre y postsináptica determinó el signo de plasticidad sináptica, con LTP resultante cuando la neurona presináptica disparó antes que la postsináptica, mientras que LTD resultó lo contrario. Esta plasticidad dependiente del tiempo de pico (STDP) fue muy emocionante porque encaja con el papel causal de la neurona presináptica en la conducción de la postsináptica. Si una determinada pre neurona realmente jugó un papel en impulsar a la postneurona al fuego, entonces necesariamente tendrá que haber disparado antes de ello, y de acuerdo con los resultados de STDP, sus pesos aumentarán de fuerza. En tanto, las pre neuronas que no tienen papel causal en el disparo de la célula postsináptica tendrán sus pesos disminuidos. Sin embargo, como discutimos con más detalle en STDP, este patrón STDP no generaliza bien a trenes de picos realistas, donde las neuronas están constantemente disparando e interactuando entre sí durante 100 milisegundos. Sin embargo, los datos de STDP proporcionan una prueba rigurosa útil para modelos computacionales de plasticidad sináptica. Basamos nuestras ecuaciones de aprendizaje en un modelo detallado utilizando mecanismos de plasticidad sináptica más básicos y con base biológica que captan estos hallazgos de STDP (Urakubo, Honda, Froemke, & Kuroda, 2008), pero que sin embargo resultan en ecuaciones de aprendizaje bastante simples cuando se consideran a nivel de velocidad de disparo.