
10.4: Detalles del procesamiento de ARNm eucariota
- Page ID
- 54138

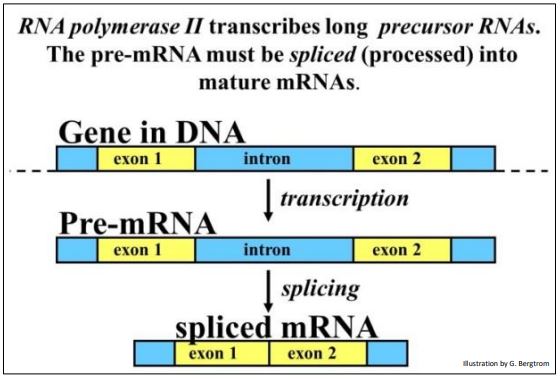
Los transcritos primarios de ARNm eucariotas se someten a un procesamiento extenso, incluyendo corte y empalme, caperuza y poliadenilación. Los pasos descritos aquí se consideran en orden de (¡a veces superpuestos!) ocurrencia. Comenzamos con el splicing, un fenómeno de mRNA.
A. Intrones empalmeosómicos
Las regiones codificantes de genes bacterianos son continuas. El descubrimiento de genes divididos eucariotas con intrones y exones fue una gran sorpresa. No solo parecía incongruente que la evolución hubiera pegado ADN irrelevante en medio del ADN codificante, ¡nadie podría haber soñado tal cosa! Por su descubrimiento de genes divididos, de Richard J. Roberts y Phillip A. Sharp compartieron el Premio Nobel de Fisiología en 1993. De hecho, todos menos algunos genes eucariotas están divididos, y algunos tienen uno, dos (¡o más de 30-50!) intrones que separan bits de ADN codificante, los exones. El empalme se resume a continuación.
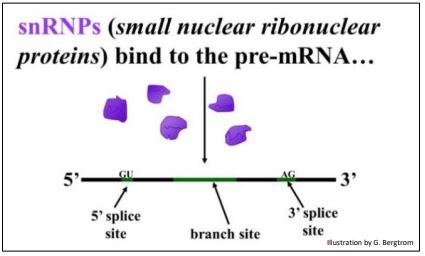
El empalme implica una serie de pequeñas proteínas ribonucleares (snRNP). Los snRNP son partículas compuestas por ARN y proteínas. Se unen a sitios específicos en un ARNm y luego dirigen una serie secuencial de cortes y ligaciones (el corte y empalme) necesarios para procesar los ARNm.
El papel de los snRNP en el corte y empalme de pre-ARNm se ilustra a continuación.
La unión de snRNP a un par de sitios de corte y empalme que flanquean un intrón en un pre-ARNm forma el spliceosoma que completa el corte y empalme, incluyendo la eliminación del lariat (la estructura intermedia del intrón). El último paso es ligar exones en un ARNm continuo con todos sus codones intactos y listos para su traducción. La acción del spliceosoma se resume a continuación.
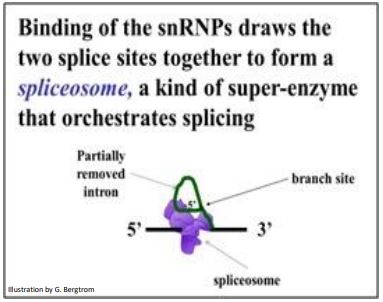
194 El descubrimiento de genes divididos
B. Cuerpos Nucleares específicos y sus proteínas asociadas facilitan el ensamblaje y la función de los SNRNP
Recordemos la organización de los núcleos facilitados por los cuerpos nucleares. Los cuerpos cajal (CB) y las gemas son cuerpos nucleares que son similares en tamaño y tienen funciones relacionadas en el ensamblaje de SNRNP spliceosómicos. Algunos defectos de corte y empalme se correlacionan con mutaciones en la proteína de la bobina que se asocian con cuerpos Cajal; otros se correlacionan con mutaciones en proteínas SMN normalmente asociadas con Gems. Una hipótesis fue que los CBs y las gemas interactúan en el ensamblaje de snRNP y spliceosoma..., pero ¿cómo? Considerar los resultados de un experimento en el que los anticuerpos contra la coilina y la proteína SMN se localizaron en células de neuroblastoma indiferenciadas y diferenciadas.
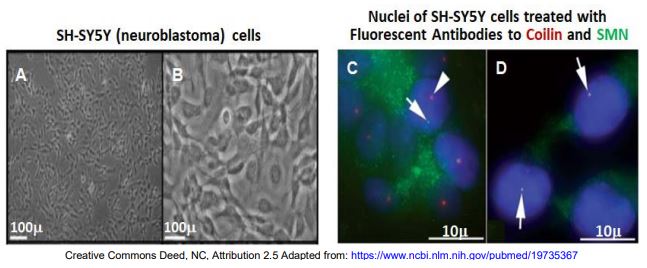
A y C son células indiferenciadas en cultivo; B y D son células que fueron estimuladas para diferenciarse. En las micrografías de fluorescencia de la derecha, las flechas apuntan a cuerpos nucleares fluorescentes. La proteína coilina se asocia con CBs y SMN se encuentra en Gems. Por lo tanto, esperamos que los anticuerpos fluorescentes contra la coilina (verde) se localicen a los CB y los anticuerpos contra la proteína SMN (rojo) se unan a Gems. Esto es lo que sucede en los núcleos de células indiferenciadas (panel C). Pero en el panel D, los dos anticuerpos se colocalizan, sugiriendo que los CB y Gems se agregan en las células diferenciadas. Esto explicaría la necesidad tanto de coilina funcional como de proteína SMN para producir snRNP funcionales. Los CB y Gems pueden estar agregándose en células diferenciadas debido a un aumento observado en la expresión de la proteína SMN. Esto podría llevar a Gemas más activas más capaces de asociarse con los CBs.
Este y otros experimentos similares demuestran que los diferentes cuerpos nucleares sí tienen funciones específicas. No son artefactos estructurales aleatorios, han evolucionado para organizar las actividades nucleares en el tiempo y el espacio de formas que son esenciales para la célula.
C. Intrones autoempalmantes del Grupo I y del Grupo II
Mientras que los intrones espliceosómicos eucariotas se empalman usando snRNP como se describió anteriormente, los intrones del Grupo I o Grupo II se eliminan por diferentes mecanismos. Los intrones del grupo I interrumpen los genes de ARNm y ARNt en bacterias y en genes mitocondriales y cloroplastos. Ocasionalmente se encuentran en genes bacteriófagos, pero raramente en genes nucleares, y luego solo en eucariotas inferiores. ¡Los intrones del Grupo I son autoempalmados! Así, son ribozimas que no requieren snRNP u otras proteínas. En cambio, se pliegan en una estructura secundaria de tallo-bucle que posiciona los nucleótidos catalíticos en sitios de corte y empalme apropiados, se extirpan y vuelven a ligar los exones. Los intrones del Grupo II en ARNr cloroplasto y mitocondrial, ARNm, ARNt y algunos ARNm bacterianos pueden ser bastante largos, formar estructuras terciarias complejas de tallo-bucle, y auto-empalme, ¡al menos en un tubo de ensayo! Sin embargo, los intrones del Grupo II codifican proteínas requeridas para su propio corte y empalme in vivo. Al igual que los intrones spliceosómicos, forman una estructura de lariato en un sitio de ramificación de residuo A. Todo esto sugiere que el mecanismo de empalme de intrones spliceosómicos evolucionó a partir del de los intrones del Grupo II.
D. Entonces, ¿por qué empalmar?
El rompecabezas que implica la pregunta, por supuesto, es por qué los organismos superiores tienen genes divididos en primer lugar. Si bien la siguiente discusión puede aplicarse a todos los empalmes, hará referencia principalmente a intrones spliceosómicos. Aquí hay algunas respuestas a la pregunta “¿Por qué empalmar?”
- Los intrones en los genes nucleares suelen ser más largos (¡a menudo mucho más largos!) que los exones. Al ser no codificantes, son grandes dianas para la mutación. En efecto, el ADN no codificante, incluidos los intrones, puede amortiguar los efectos nocivos de las mutaciones aleatorias.
- Puede recordar que la duplicación de genes en un cromosoma (y la pérdida de una copia de su homólogo) surgen de recombinación desigual (cruce no homólogo). Ocurre cuando secuencias de ADN similares se alinean durante la sinapsis de meiosis. En un organismo que hereda un cromosoma con ambas copias génicas, el duplicado puede acumular mutaciones siempre y cuando el otro conserve la función original. El gen divergente se convierte entonces en parte de un acervo de ADN seleccionable, el grano de la evolución, en los descendientes de organismos que heredan los genes duplicados, aumentando la diversidad de especies. La recombinación desigual también puede ocurrir entre secuencias similares (por ejemplo, en intrones) en los mismos o diferentes genes. Los intrones también pueden permitir el intercambio de exones entre genes. Después de una recombinación desigual entre intrones que flanquean un exón, un gen adquirirá otro exón mientras que el otro lo perderá. Una vez más, siempre y cuando un organismo conserve una copia de los genes participantes con función original, el organismo puede producir la proteína requerida y sobrevivir. En tanto, el gen con el exón extra puede producir la misma proteína, pero una con un nuevo dominio estructural y función. Al igual que un gen duplicado completo, uno con un nuevo exón y función añadida está en el acervo de ADN seleccionable. Así, ¡este fenómeno de barajado de exones aumenta la diversidad de especies! La evidencia indica que se ha producido el barajado de exones, creando proteínas con diferentes funciones globales que, sin embargo, comparten al menos un dominio y una función común. Un ejemplo discutido anteriormente involucra proteínas de unión a calcio que regulan muchos procesos celulares. Los dominios de unión a calcio (Ca++) relacionados estructuralmente son comunes a muchas proteínas estructuralmente y funcionalmente no relacionadas. Considere el barajado de exones en el cruce desigual (recombinación no homóloga) que se ilustra a continuación.
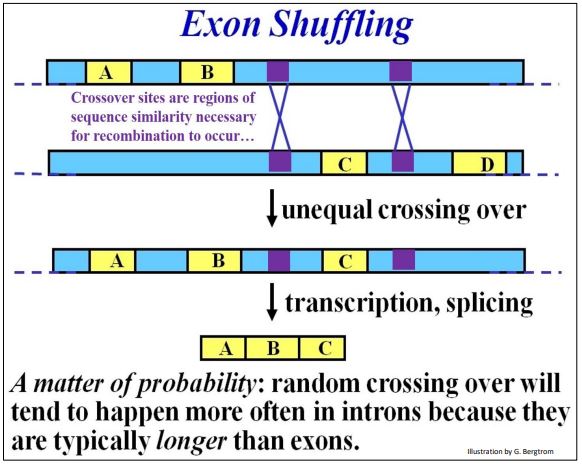
En este ejemplo, regiones de fuerte similitud en diferentes intrones (no homólogos) en el mismo gen se alinean durante la sinapsis de meiosis. El cruce desigual entre los genes inserta el exón C en uno de los genes. El otro gen pierde el exón (no mostrado en la ilustración). En suma, los intrones son tampones contra mutaciones deletéreas, e igualmente valiosos, son dianas potenciales para la duplicación de genes y el barajado de exones. Esto convierte a los intrones en actores clave en la creación de diversidad genética, el sello distintivo de la evolución.
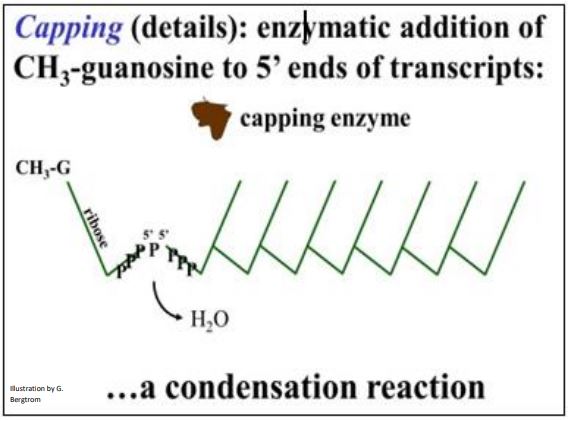
E. Tapado
Una tapa de metil guanosina añadida de 5' a 5' a un ARNm funciona en parte para ayudar a los ARNm a abandonar el núcleo y asociarse con ribosomas. La tapa se agrega a un extremo 5' expuesto, incluso cuando la transcripción y el corte y empalme aún están en progreso. Una enzima de caperuza coloca un residuo de guanosina metilado en el extremo 5' del ARNm maduro. La estructura de tapa 5' se muestra a continuación (las marcas de verificación son nucleótidos unidos 5'-3').
F. Poliadenilación
Después de la terminación de la transcripción, la poli (A) polimerasa cataliza la adición de múltiples residuos AMP (varios cientos en algunos casos) al extremo 3' por la enzima. La enzima se une a una secuencia de AAUAA cerca del extremo 3' de un ARNm y comienza a catalizar la adición de los monofosfatos de adenosina. El sitio de reconocimiento de poli (A) de AAUAA se indica en rojo en la ilustración de poliadenilación que se muestra a continuación.
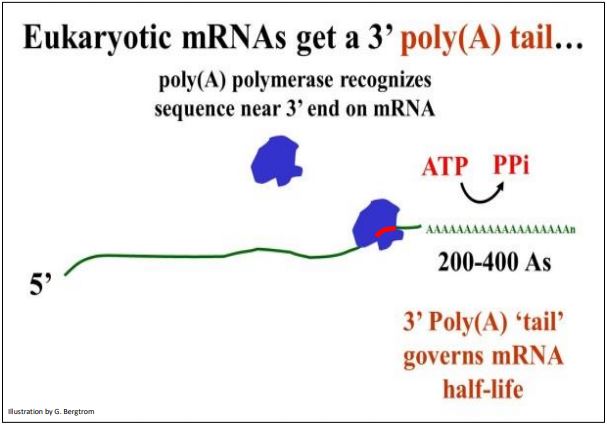
El resultado de la poliadenilación es una cola poli (A) 3' cuyas funciones incluyen ayudar en el tránsito de los ARNm desde el núcleo y regular la vida media de los ARNm en el citoplasma. La cola poli (A) se acorta cada vez que un ribosoma completa la traducción del ARNm.