
10.5: Los ARNt son patos bastante extraños
- Page ID
- 53368

En procariotas, el ARNt se puede encontrar ya sea como genes individuales o como partes de operones que también pueden contener combinaciones de ARNm o ARNr. En cualquier caso, ya sea de un solo gen, o después de la escisión inicial para separar el transcrito de ARNt del resto del transcrito, el pre-ARNt resultante tiene un líder N-terminal (41 nt en E. coli) que es escindido por RNasa P. Esa escisión es universal para cualquier ARNt procariota. Después de eso, hay variaciones en las escisiones menores realizadas por una variedad de nucleasas que producen el ARNt en su longitud final aunque no en su secuencia madura, como veremos en algunos párrafos.
El pre-ARNt eucariota (transcrito por la ARN polimerasa III) de manera similar tiene un líder N-terminal eliminado por la RNasa P. Sin embargo, a diferencia del procariota, la longitud puede variar entre diferentes ARNt de la misma especie. Algunos transcritos de pre-ARNt eucariotas también contienen intrones, especialmente en el bucle anticodón, que deben ser cortados y empalmados para que el ARNt funcione normalmente. Estos intrones son diferentes de los transcritos autoempalmados o spliceosoma-empalmados discutidos en el capítulo de transcripción. Aquí, la función de corte y empalme se lleva a cabo no por ribozimas, sino por enzimas convencionales (proteínicas). Curiosamente, RNaseP también elimina una secuencia 3' del pre-ARNt, pero luego se agrega otra secuencia 3' nuevamente. Este nuevo extremo 3' es siempre CCA, y se agrega por tres rondas sucesivas con ARNt nucleotidiltransferasa.
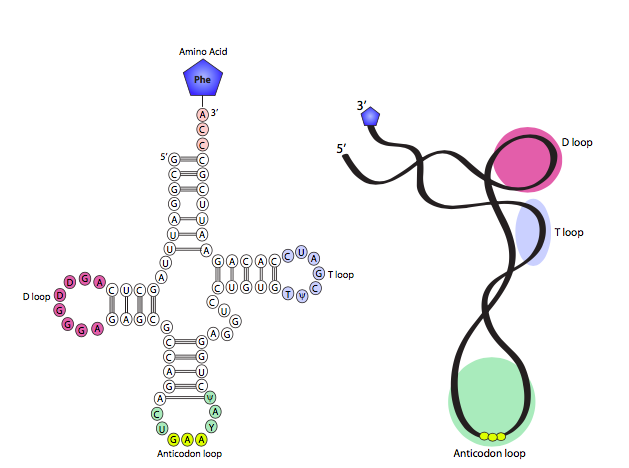
Cuando se introdujo el ARN, se observó que aunque extremadamente similar al ADN en muchos aspectos, normalmente es monocatenario, y esa propiedad, combinada con la oportunidad de apareamiento de bases complementarias dentro de una cadena, le permite hacer algo muy diferente al ADN bicatenario: puede formar altamente complejo estructuras secundarias. Uno de los ejemplos más simples y claros de esto es el ARNt, que depende de su conformación para lograr su función celular. El diagrama prototípico de ARNt en forma de hoja de trébol se muestra en la Figura\(\PageIndex{2}\) de la izquierda, con un modelo 3D derivado de datos cristalográficos de rayos X a la derecha. Como puede ver, la forma completamente expandida tiene cuatro “brazos” tallo-bucle con el aminoácido unido al brazo aceptor, que está en el lado opuesto del ARNt del brazo anticodón, que es donde el ARNt debe coincidir con el codón de ARNm durante la traducción. Aproximadamente perpendiculares al eje aceptor-anticodón están el brazo D y el brazo TyC. En algunos ARNt, en realidad hay cinco brazos totales con un bucle muy corto entre los brazos TyC y anticodón. Las estructuras de tallo y bucle en forma de brazo están formadas por dos áreas de fuerte complementariedad (los tallos, pares de bases) interrumpidas por una secuencia corta no complementaria (el bucle). En términos generales, los brazos se utilizan para posicionar adecuadamente el ARNt dentro del ribosoma, así como reconocer el codón de ARNm y traer el aminoácido correcto.
Cuando llega el momento de que el ARNt haga coincidir su anticodón con el codón en el ARNm, el código no se sigue “al pie de la letra” si vas a perdonar el juego de palabras. Existe un fenómeno llamado “bamboleo” en el que se permite una coincidencia codon-anticodón y se estabiliza para la traducción aunque el nucleótido en la tercera posición no sea complementario. La oscilación puede ocurrir porque la conformación del ARNt permite un poco de flexibilidad a esa posición del anticodón, permitiendo que se formen enlaces H donde normalmente no lo harían. Sin embargo, este no es un fenómeno universal: solo se aplica a situaciones en las que una U o una G se encuentra en la primera posición del anticodón (coincidiendo con la tercera posición del codón). Siguiendo la convención de secuencias de ácidos nucleicos, la secuencia siempre se escribe 5' a 3', aunque en el caso de la coincidencia codon-anticodón, las cadenas de ARNm y ARNt son antiparalelas:
Además de permitir un poco de bamboleo en el emparejamiento de bases complementarias, las moléculas de ARNt tienen otra peculiaridad. Después de incorporarse inicialmente a un ARNt a través de la transcripción convencional, existe una modificación extensa de algunas de las bases del ARNt. Esto afecta tanto a las purinas como a las pirimidinas, y puede ir desde simples adiciones como la metilación o la reestructuración extensa del propio esqueleto del azúcar, como en la conversión de guanosina en wyosina (W). Hasta la fecha se han catalogado más de 50 modificaciones diferentes. Estas modificaciones pueden ser casi universales, como la dihidrouridina (D) que se encuentra en el bucle D del ARNt, o más específicas, como la conversión de G a W que se encuentra principalmente en ARNtPHE de ciertas especies (se han identificado ejemplos tanto en especies procariotas como eucariotas). Hasta el 10% de las bases en un ARNt pueden ser modificadas. Naturalmente, las alteraciones en la base de azúcar de los nucleótidos también pueden alterar las características de apareamiento de bases. Por ejemplo, una base modificada común, la inosina, puede complementar U, C o A. Este apareamiento aberrante de bases complementarias puede ser igual entre las bases pretendientes, o puede estar sesgado, como en el caso de la 5-metoxiuridina, que puede reconocer A, G o U, pero el reconocimiento de U es pobre.
Cargar el ARNt
El conocimiento del código genético plantea la pregunta: ¿cómo se une el aminoácido correcto a algún ARNt dado? Una clase de enzimas llamadas aminoacil ARNt sintetasas son las encargadas de reconocer tanto un ARNt específico como un aminoácido específico, uniendo un ATP para obtener energía y luego unirlos (a veces llamados “cargar el ARNt”) con hidrólisis del ATP. La especificidad es una tarea difícil para la sintetasa ya que los aminoácidos se construyen a partir del mismo esqueleto y son muy similares en masa. Distinguir entre moléculas de ARNt es más fácil, ya que son más grandes y sus estructuras secundarias también permiten una mayor variación y por lo tanto mayor facilidad de discriminación. También existe un mecanismo de corrección de preadhesión incorporado en el que las moléculas de ARNt que se ajustan bien a la sintetasa (es decir, las correctas) mantienen el contacto por más tiempo y permiten que la reacción continúe, mientras que las moléculas de ARNt mal ajustadas e incorrectas probablemente se desasocien de la sintetasa antes de que intente adjuntar el aminoácido.
Cargar una aminoacil ARNt sintetasa con su aminoácido requiere energía. La sintetasa se une primero a una molécula de ATP y al aminoácido apropiado, los cuales reaccionan dando como resultado la formación de aminoacil-adenilato y pirofosfato. El PP i se libera y la sintetasa ahora se une al ARNt apropiado. Finalmente, el aminoácido se transfiere al ARNt. Dependiendo de la clase de sintetasa, el aminoácido puede unirse al 2'-OH del terminal A (clase I) o al 3'-OH del terminal A (clase II) del ARNt. La Phe-ARNt sintetasa es la excepción: es estructuralmente una enzima de clase II pero transfiere el Phe al 2'-OH. Obsérvese que los aminoácidos transferidos al 2'-OH pronto se mueven al 3'-OH de todos modos debido a una reacción de transesterificación.