
12.3: Microfilamentos de Actina
- Page ID
- 53333

Los microfilamentos también se conocen como filamentos de actina, actina filamentosa y f-actina, y son los opuestos citoesqueléticos de los filamentos intermedios. Estas hebras están compuestas por pequeñas subunidades de actina globular (g-actina) que se apilan entre sí con puntos de contacto relativamente pequeños. Podrías imaginar dos pelotas de tenis, una borrosa y la otra cubierta con ganchos de velcro. Incluso si presiona con fuerza para mezclarlas, el área de contacto entre las bolas (es decir, el área disponible para la unión H entre subunidades) es bastante pequeña en comparación con el área de superficie general, o con el área de contacto entre las subunidades IF. Ellos se mantendrán unidos, pero también pueden desmoronarse con relativamente poca fuerza. Contraste esto con filamentos intermedios, que podrían representarse como dos cintas de ganchos o bucles de velcro. Se requiere considerablemente más trabajo para desarmarlos. Debido a que hay menos enlaces H que romper, los microfilamentos se pueden deconstruir muy rápidamente, lo que los hace adecuados para aplicaciones altamente dinámicas.
Cuando las subunidades de actina se unen para formar microfilamentos, interactúan direccionalmente. Es decir, las subunidades tienen una “parte superior” y una “parte inferior”, y la parte superior de una subunidad siempre interactúa con la parte inferior de otra. Si vamos a la subunidad más “inferior” de un filamento, el extremo abierto se llama el extremo menos (-), mientras que el extremo opuesto, que por cierto ve más acción aditiva, se llama el extremo más (+). También se dice que los microfilamentos tienen polaridad, pero nuevamente esto es sólo en el sentido de tener direccionalidad, y no tiene nada que ver con la carga eléctrica. Los microfilamentos individuales pueden existir, pero la mayoría de los microfilamentos in vivo son pares trenzados. A diferencia del ADN; sin embargo, los pares de microfilamentos no son antiparalelos: ambas hebras tienen la misma direccionalidad.
La formación de filamentos a partir de la g-actina es un proceso dependiente del ATP, aunque no en el sentido convencional de utilizar la energía liberada en la hidrólisis. En cambio, las subunidades globulares de actina solo se unirán con otra subunidad de g-actina si primero se ha unido a un ATP. Si la g-actina tiene ADP unido, entonces primero debe cambiar el ADP por ATP antes de que pueda agregarse a un filamento. Esto altera la conformación de la subunidad para permitir una interacción de mayor afinidad. Poco tiempo después, la hidrólisis del ATP a ADP (con liberación de P i) debilita la afinidad pero no causa directamente la disolución de la unión de la subunidad. La hidrólisis es producida por la actina misma, que tiene esta actividad enzimática de ATPasa incorporada.
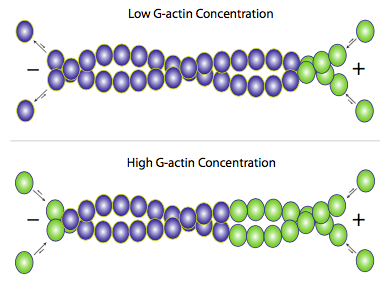
Aunque la f-actina existe principalmente como un par de filamentos retorcidos uno alrededor del otro, la adición de nueva actina ocurre mediante la adición de monómeros de g-actina individuales a cada filamento (Figura\(\PageIndex{3}\)). Las proteínas accesorias se pueden utilizar para ayudar o dificultar la construcción o descomposición de los filamentos, pero el mecanismo principal es esencialmente autorregularse. Cuando los niveles de g-actina libre son altos, se favorece el alargamiento de los filamentos de actina, y cuando la concentración de g-actina disminuye, predomina la despolimerización de la f-actina. Sin embargo, bajo condiciones fisiológicas promedio, lo que a menudo se ve en los microfilamentos de actina es un efecto llamado treadmilling. Dado que la actina se agrega principalmente en un extremo pero se elimina del otro, el efecto neto es que cualquier monómero de actina dado en un filamento se mueve efectivamente del extremo (+) al extremo (-) incluso si la longitud aparente del filamento no cambia.
En la mayoría de los tipos celulares, la mayor concentración de estructuras citoesqueléticas basadas en actina se encuentra en la periferia de la célula en lugar de hacia el centro. Esto encaja bien con la tendencia de los bordes de la célula a ser más dinámicos, ajustándose constantemente para sentir y reaccionar ante su entorno. Claramente, la polimerización y despolimerización de los filamentos de actina es mucho más rápida que para los filamentos intermedios. La gran excepción a la regla de actina en la periferia se encuentra en las células musculares. Los filamentos de actina, y las proteínas motoras de miosina que trabajan en ellos, son la base para la contracción de las células musculares, y suben la mayoría de las células musculares, no solo la periferia. Discutiremos el papel de la actina en ambos tipos de movimiento celular más adelante en el capítulo.