
2.5: Estructura y Función- Función Proteína II
- Page ID
- 52942

Fuente: BiochemFFA_2_4.pdf. Todo el libro de texto está disponible de forma gratuita de los autores en http://biochem.science.oregonstate.edu/content/biochemistry-free-and-easy
A este punto, las proteínas que hemos discutido no han sido catalizadores (enzimas). La mayoría de las proteínas en las células, sin embargo, catalizan reacciones. En esta sección comenzamos nuestra discusión sobre una subclase de proteínas que catalizan reacciones liberando energía y la convierten en fuerza mecánica. Estos operan a nivel celular y organizativo y se conocen como proteínas motoras. Las proteínas motoras se basan en proteínas estructurales globulares, por lo que es importante que describamos cómo se ensamblan estos “ferrocarriles” celulares antes de discutir las propias proteínas motoras. Existen dos estructuras fibrosas relevantes que sirven como rieles para las proteínas motoras. Ellos son:
- microfilamentos (compuestos de un polímero de actina) y
- microtúbulos (compuestos por un polímero de tubulina.
Actina
La unidad monomérica de actina se llama G-actina (actina globular) y el polímero se conoce como F-actina (actina filamentosa). Los filamentos de Factina comprenden los filamentos más pequeños de células conocidos como microfilamentos (Figura 2.101). La actina es esencial para la contracción muscular y también tiene diversos papeles en la señalización celular y el mantenimiento de las uniones celulares. Junto con otras proteínas, la actina tiene numerosas interacciones con la membrana celular. Las formas β y γ de la actina son componentes del citoesqueleto y facilitan la motilidad dentro de las células. La α-actina es importante en los tejidos musculares, donde es utilizada por la miosina en el proceso mecánico de contracción (Ver AQUÍ).
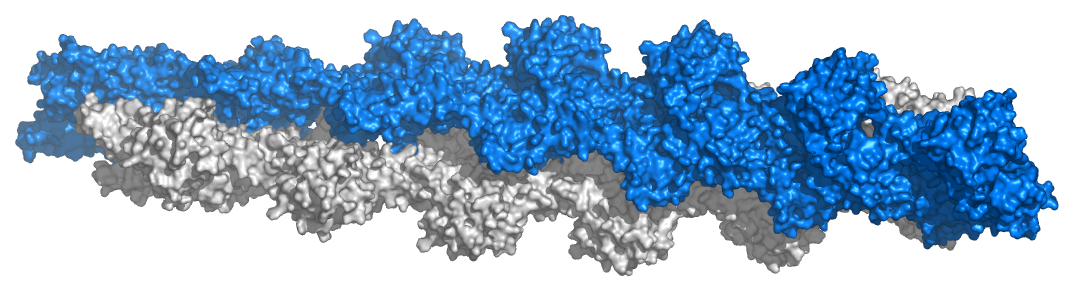
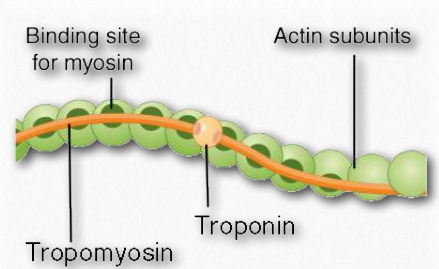
Las formas monoméricas y poliméricas de actina desempeñan un papel en las actividades celulares relacionadas con el movimiento. Dos hebras paralelas de F-actina pueden emparejarse entre sí y crear una estructura helicoidal doble con 2.17 subunidades por giro de la hélice. La F-actina helicoidal en los músculos contiene tropomiosina, que cubre los sitios de unión de actina para la miosina en los músculos en reposo para evitar la contracción. Otras proteínas unidas a los filamentos musculares de actina incluyen las troponinas (I, T y C).
Actina Acción Celular
Los ejemplos de acción de actina a nivel celular incluyen motilidad celular, citocinesis, transporte intracelular de vesículas y orgánulos, y forma celular. Cada monómero de actina está unido a una molécula de ATP o ADP y la presencia de uno de estos es esencial para el correcto funcionamiento de la G-actina.
El papel de ATP
En el monómero, la actina se une más comúnmente a ATP, mientras que en los filamentos, normalmente se une a ADP. La actina es una ATPasa ineficiente, descomponiendo la molécula lentamente, pero la catálisis se acelera hasta 40,000 veces cuando el monómero comienza a polimerizarse. La actina también tiene un sitio de unión para cationes divalentes, ya sea calcio o magnesio. F-
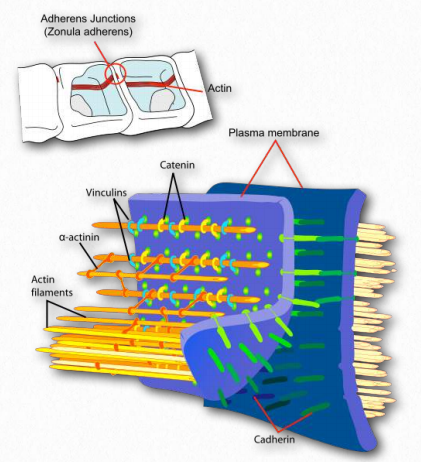
La actina se une a proteínas estructurales en la unión de adherentes (Figura 2.102). Estos incluyen α-actinina, vinculina (proporciona una conexión de membrana y conexiones a las cateninas y cadherina).
Polimación
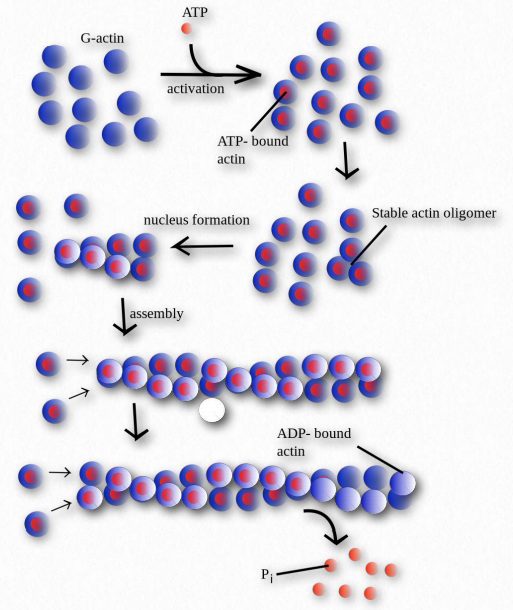
La polimerización de actina comienza con un evento nucleante (Figura 2.103). Un factor que se sabe que afecta el proceso se conoce como el complejo Arp 2/3. Lo hace imitando un dímero de actina, iniciando un proceso autocatalítico de ensamblaje de actina. El complejo Arp 2/3 juega un papel tanto en el inicio de la polimerización de nuevos filamentos de actina como en la formación de ramas en los filamentos.
Dos proteínas juegan un papel en la modulación del crecimiento de polímeros. La timosina funciona en el extremo de los filamentos de actina para controlar el crecimiento. Profilin trabaja en monómeros de G-actina intercambiando ADP por ATP, promoviendo la adición de monómeros a una cadena en crecimiento.
Los filamentos de F-actina se mantienen unidos por enlaces relativamente débiles en comparación con los enlaces covalentes de los monómeros de ácidos nucleicos, permitiendo así un desmontaje más fácil cuando se desee. La secuencia de aminoácidos de actina está optimizada, habiendo divergido solo una cantidad relativamente pequeña (20%) entre algas y humanos. Las mutaciones en el gen de actina resultan en enfermedades musculares y/o sordera.
Tubulina
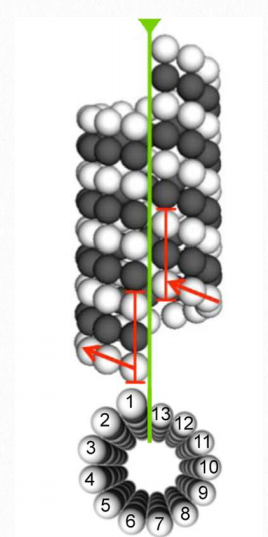
Las proteínas tubulina son los bloques de construcción monoméricos de los microtúbulos eucariotas (Figura 2.104 y 2.105). Se conocen equivalentes bacterianos (TuBZ) y arqueones (FtsZ). Las proteínas α-tubulina y β-tubulina se polimerizan para hacer estructuras de microtúbulos en el citoplasma de las células. Los microtúbulos son componentes principales del citoesqueleto de las células eucariotas, proporcionando soporte estructural, transporte dentro de la célula y funciones necesarias para la segregación de ADN durante la división celular.
La dimerización de las proteínas α-tubulina y β-tubulina es necesaria para la polimerización y requiere que las subunidades se unan a GTP. Los microtúbulos solo crecen en una dirección. β- tubulina se encuentra en el extremo más del túbulo (extremo de crecimiento = extremo más) y α-tubulina se expone en el otro extremo (extremo sin crecimiento = extremo menos). Los dímeros de α-tubulina/β-tubulina se incorporan en los microtúbulos en crecimiento en esta orientación. Si un dímero está ligado al PIB en lugar de GTP, tiende a ser inestable y desmoronarse, mientras que los unidos a GTP se ensamblan establemente en microtúbulos.
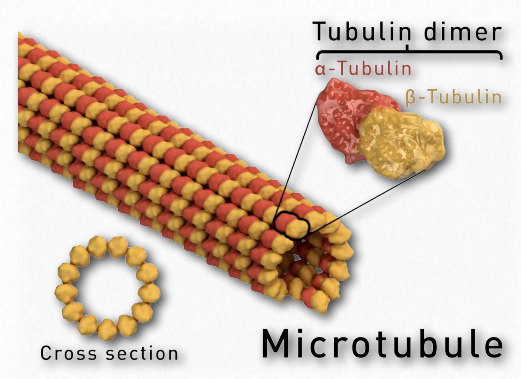
Microtúbulos
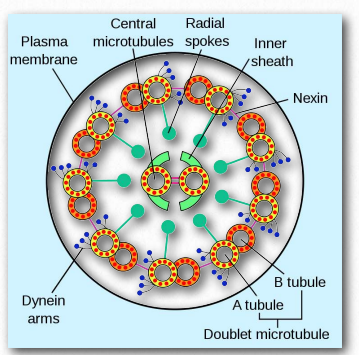
Los microtúbulos, junto con los microfilamentos y los filamentos intermedios (ver AQUÍ) constituyen el citoesqueleto de las células. Se encuentran en el citoplasma, se encuentran en células eucariotas, así como en algunas bacterias. Los microtúbulos ayudan a dar estructura a las células. Comprenden la estructura interna de flagelos y cilios y proporcionan superficies similares a rieles para el transporte de materiales dentro de las celdas.
La polimerización de α- tubulina y β-tubulina para formar microtúbulos ocurre después de un evento nucleante. Las unidades individuales se organizan en centros de organización de microtúbulos (MTOC), un ejemplo de lo cual es el centrosoma. Los centrosomas son puntos focales de conexión de microtúbulos. Los cuerpos basales de cilios y flagelos también ayudan a organizar los microtúbulos.
Proteínas motoras
Desde el transporte de materiales dentro de una célula hasta el proceso de citocinesis donde una célula se divide en dos en la mitosis, una célula tiene necesidades de movimiento a nivel molecular. Deben transportarse vesículas secretoras y orgánulos. Los cromosomas deben separarse en mitosis y meiosis.
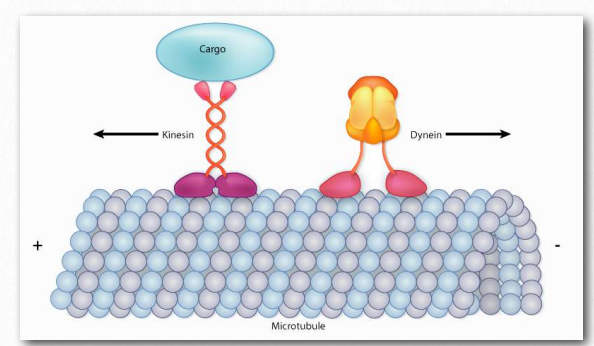
Las proteínas dineína y cinesina (Figura 2.106) son necesarias para el movimiento intracelular. Estas proteínas motoras facilitan el movimiento de materiales dentro de las células a lo largo de “rieles” de microtúbulos. Estas proteínas motoras son capaces de moverse a lo largo de una porción del citoesqueleto al convertir la energía química en movimiento con la hidrólisis del ATP. Una excepción es la rotación flagelar, que utiliza la energía proporcionada por un gradiente creado por una bomba de protones.
Kinesinas y dineínas
Como se señaló, las quinesinas y las dineinas navegan en células en pistas de microtúbulos (Figura 2.108 y Película 2.4). La mayoría de las quinesinas se mueven en la dirección de la síntesis del microtúbulo (+ movimiento final), que generalmente se aleja del centro celular y la dirección opuesta de movimiento de las dineinas, que se dice que hacen transporte retrógrado hacia el centro celular. Ambas proteínas proporcionan funciones de movimiento necesarias para los procesos de mitosis y meiosis. Estos incluyen la formación de huso, la separación de cromosomas y el transporte de orgánulos, como las mitocondrias, los aparatos de Golgi y las vesículas.
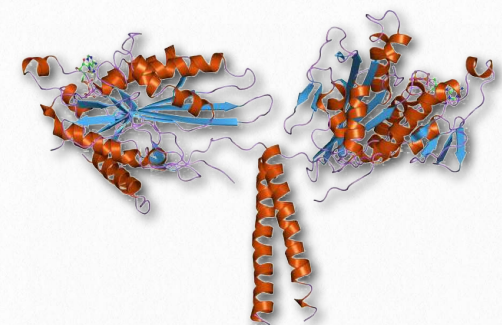
Las quinesinas están compuestas por dos cadenas pesadas y dos cadenas ligeras. Los dominios motores de cabeza de las cadenas pesadas (en los pies) utilizan la energía de la hidrólisis de ATP para realizar trabajos mecánicos para el movimiento a lo largo de los microtúbulos. Hay al menos catorce familias distintas de cinesina y probablemente muchas familias relacionadas además.
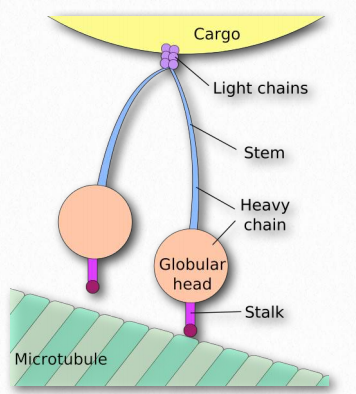
Las dineínas se colocan en dos grupos: citoplasmático y axonémico (también llamadas dineínas ciliares o flagelares - Figura 2.109). Las dineínas son de estructura más compleja que las quinesinas con muchas pequeñas unidades polipeptídicas. Notablemente, las plantas no tienen proteínas motoras de dineína, pero sí contienen quinesinas.
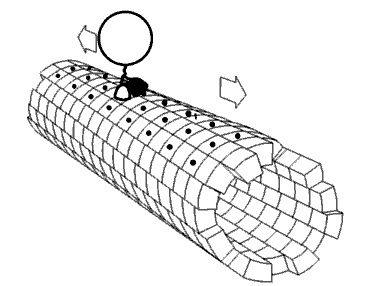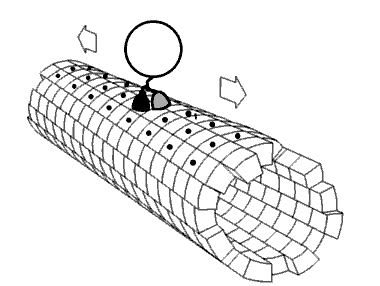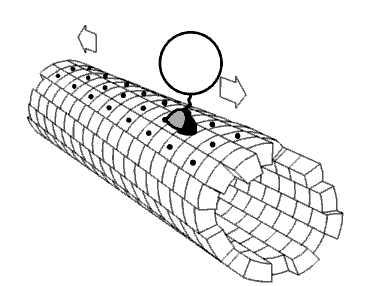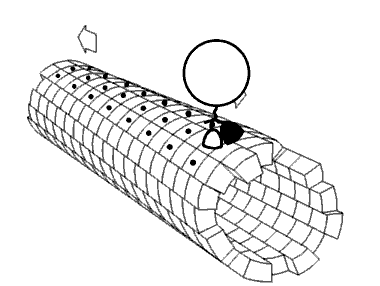
Película 2.4 La cinesina de la proteína motora caminando por un microtúbulo. Imagen utilizada con permiso (Dominio Público; zp706).
Miosina
Un grupo importante de proteínas motoras en la célula son las miofinas. Al igual que las quinesinas y las dineinas, las miosas utilizan la energía de la hidrólisis del ATP para el movimiento. En este caso, el movimiento no es principalmente a lo largo de microtúbulos, sino a lo largo de microfilamentos compuestos por un polímero de actina (F-actina). El movimiento de la miosina sobre la actina es mejor conocido como la fuerza impulsora de la contracción muscular. Las miosinas son una enorme familia de proteínas, todas las cuales se unen a la actina y todas ellas implican movimiento. Se conocen dieciocho clases diferentes de proteínas miosina.
La miosina II es la forma responsable de generar contracción muscular. Es una proteína alargada formada a partir de dos cadenas pesadas con cabezas motoras y dos cadenas ligeras. Cada cabeza motora de miosina se une a actina y tiene un sitio de unión a ATP. Las cabezas de miosina se unen e hidrolizan ATP. Esta hidrólisis produce la energía necesaria para que la miosina camine hacia el extremo positivo de un filamento de actina.
La miosina II no muscular proporciona la contracción necesaria para potenciar la acción de la citocinesis. Otras proteínas miosina están involucradas en el movimiento de las células no musculares. La miosina I está involucrada en la organización intracelular. La miosina V realiza el transporte de vesículas y orgánulos. La miosina XI proporciona movimiento a lo largo de las redes de microfilamentos celulares para facilitar el flujo de orgánulos y citoplásmicos en una dirección particular.
Estructura
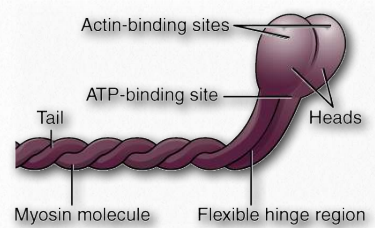
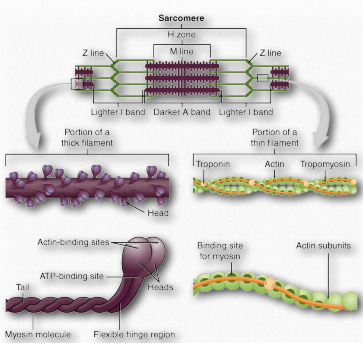
Las miosinas tienen seis subunidades, dos cadenas pesadas y cuatro cadenas ligeras. Las proteínas miosina tienen dominios frecuentemente descritos como cabeza y cola (Figura 2.111). Algunos también describen una región bisagra intermedia como cuello. La porción de cabeza de la miosina es la parte que se une a la actina. Utiliza energía de la hidrólisis de ATP para moverse a lo largo de los filamentos de actina. En los músculos, las proteínas miosina forman estructuras agregadas denominadas filamentos gruesos. Los movimientos son direccionales.
Consideraciones estructurales de la contracción muscular
Antes de discutir los pasos en el proceso de contracción muscular, es importante describir los aspectos anatómicos de los músculos y la nomenclatura.
Hay tres tipos de tejido muscular: esquelético (estriado), liso y (en vertebrados) cardiaco. Nos ocuparemos principalmente aquí del tejido muscular esquelético. Los músculos pueden ser activados por el sistema nervioso central o, en el caso de los músculos lisos y cardíacos, pueden contraerse involuntariamente. Los músculos esqueléticos pueden ser contracción lenta o contracción rápida.
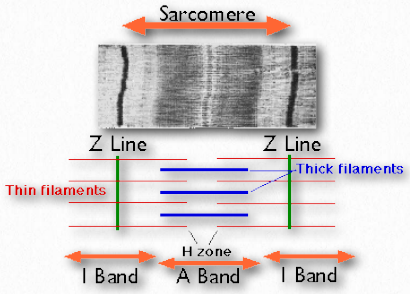
Sarcómeros
Los sarcómeros se describen como las unidades básicas que comprenden músculos estriados y están compuestos por filamentos gruesos (miosina) y delgados (actina) y una proteína llamada titina. Los filamentos se deslizan uno junto al otro en contracción muscular y luego hacia atrás en relajación muscular. No se encuentran en los músculos lisos.
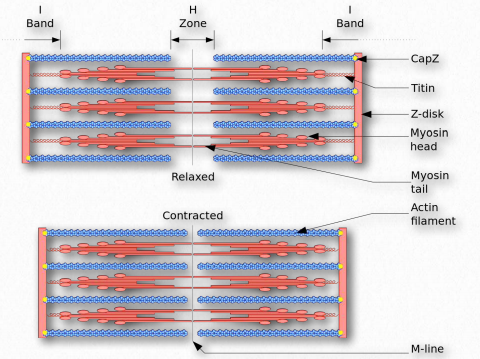
Bajo el microscopio, un sarcómero es la región entre dos líneas Z de tejido muscular estriado (Figura 2.112). La línea Z es la región clara, estrecha y oscura en el medio de una banda I. Dentro del sarcómero se encuentra todo un Aband con su zona H central. Dentro de la zona Hzone se encuentran colas de fibras de miosina, con la cabeza apuntando hacia afuera desde allí proyectándose hasta la banda I. El exterior del Aband es el más oscuro y se vuelve más ligero moviéndose hacia el centro.
Dentro de la Iband se localizan filamentos delgados que no están ocupados con filamentos gruesos de miosina. El Aband contiene filamentos gruesos intactos superpuestos a filamentos delgados excepto en la zona H central, que contiene solo filamentos gruesos. En el centro de la zona H se encuentra una línea, conocida como la línea M. Contiene elementos de conexión del citoesqueleto celular. En la contracción muscular, las cabezas de miosina caminan tirando de sus colas sobre los filamentos delgados de actina, utilizando la energía de la hidrólisis del ATP y tirando de ellos hacia el centro del sarcómero.
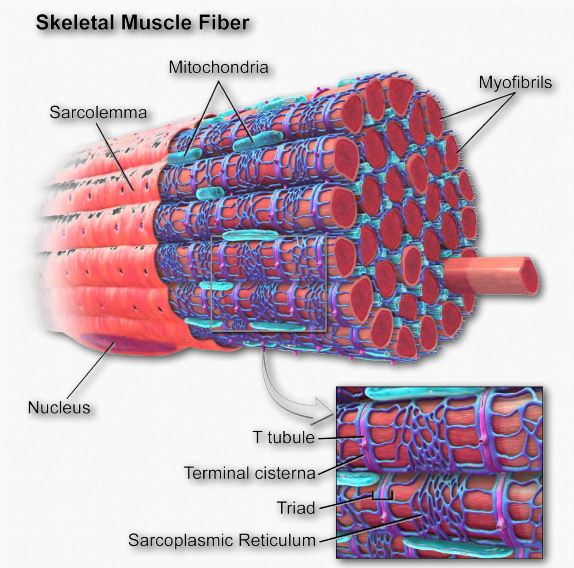
Sarcolemma
El sarcolema (también conocido como el miolema) es para las células musculares lo que es la membrana plasmática para otras células eucariotas, una barrera entre el interior y el exterior. Contiene una bicapa lipídica y un glicocáliz en el exterior de la misma. El gliocálix contiene polisacáridos y se conecta con la membrana basal. La membrana basal sirve un andamio s para conectar las fibras musculares a. Esta conexión se realiza mediante proteínas transmembrana que unen el citoesqueleto de actina en el interior de la célula con la membrana basal en el exterior. En los extremos de las fibras musculares, cada sarcolema se fusiona con una fibra tendinosa y estas, a su vez, se adhieren a los huesos.
Retículo sarcoplásmico
El retículo sarcoplásmico (Figura 2.114) es un nombre para la estructura que se encuentra dentro de las células musculares que es similar al retículo endoplásmico liso que se encuentra en otras células. Contiene un conjunto especializado de proteínas para satisfacer necesidades únicas de las células musculares. El orgánulo sirve en gran medida como una “batería” de calcio, liberando calcio almacenado para iniciar la contracción muscular cuando se estimula y absorbiendo calcio cuando se señala al final del ciclo de contracción. Realiza estas tareas utilizando canales de iones de calcio para la liberación del ion y bombas específicas de iones de calcio para absorberlo.
Dirección del movimiento
Todas las miosina excepto la miosina VI se mueven hacia el extremo + (el extremo creciente) del microfilamento. La porción del cuello sirve para unir la cabeza y la cola. También es un sitio de unión para proteínas de cadena ligera de miosina que forman parte de un complejo macromolecular con funciones reguladoras. La cola es el punto de unión de moléculas u otra “carga” que se está moviendo. También puede conectarse con otras subunidades de miosina y puede tener un papel que desempeñar en el control del movimiento.
Contracción muscular
El modelo de filamento deslizante se ha propuesto para describir el proceso de tensión/contracción muscular. En este proceso, un conjunto repetitivo de acciones desliza un delgado filamento de actina sobre un filamento grueso de miosina como medio para crear tensión/acortamiento de la fibra muscular.
Los pasos en el proceso ocurren de la siguiente manera:
A. Una señal del sistema nervioso central (potencial de acción) llega a una neurona motora, que transmite hacia la unión neuromuscular (ver más sobre la parte de neurotransmisión del proceso AQUÍ)
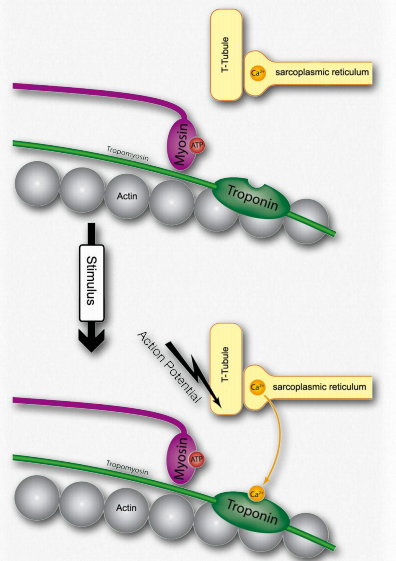
B. Al final del axón, la señal nerviosa estimula la apertura de los canales de calcio en el extremo axón haciendo que el calcio fluya hacia el terminal.
C. El movimiento del calcio hacia el axón del nervio provoca que la acetilcolina (neurotransmisor) en las vesículas sinápticas se fusione con la membrana plasmática. Esto hace que la acetilcolina sea expulsada a la hendidura sináptica entre el axón y la fibra muscular esquelética adyacente.
D. La acetilcolina se difunde a través de la sinapsis y luego se une a los receptores nicotínicos de acetilcolina en la unión neuromuscular, activándolos.
E. La activación del receptor estimula la apertura de puertas de los canales de sodio y potasio, permitiendo que el sodio se mueva hacia la célula y el potasio salga. La polaridad de la membrana de la célula muscular (llamada sarcolema - Figura 2.111) cambia rápidamente (llamada potencial de placa terminal).
F. El cambio en el potencial de la placa final da como resultado la apertura de canales iónicos sensibles al voltaje específicos para sodio o potasio solo a la Figura 2.117 - 3. La escisión de ATP por miosina permite la unión de actina (J) Wikipediaopen, creando un potencial de acción (cambio de voltaje) que se extiende por toda la célula en todas las direcciones.
G. El potencial de acción de propagación despolariza la fibra muscular interna y abre los canales de calcio en el retículo sarcoplásmico (Figura 2.115).
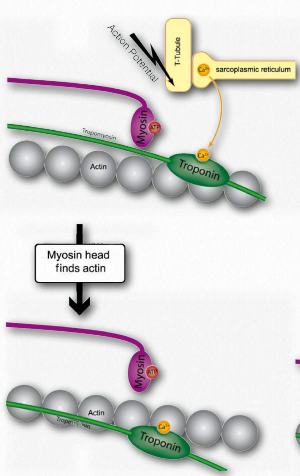
H. El calcio liberado del retículo sarcoplásmico se une a la troponina en los filamentos de actina (Figura 2.115).
I. La troponina altera la estructura de la tropomiosina a la que está unida. Esto hace que la tropomiosina se mueva ligeramente, permitiendo el acceso a los sitios de unión a miosina en el microfilamento (también llamado filamento delgado) que estaba cubriendo (Figura 2.116).
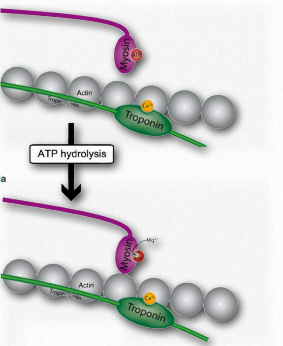
J. La miosina (unida a ATP) escinde el ATP a ADP y Pi, que retiene en su región de cabeza y luego se une a los sitios de unión expuestos en los filamentos delgados provocando que el fosfato inorgánico se libere de la miosina seguido de ADP (Figura 2.117).
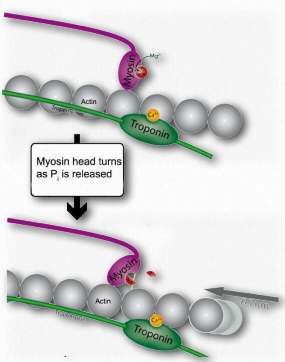
K. La liberación de ADP y Pi está estrechamente acoplada a una flexión de la bisagra de la miosina, lo que resulta en lo que se llama el golpe de potencia. Esto hace que el filamento delgado se mueva con relación a las fibras gruesas de la miosina (Figuras 2.118 y 2.119).
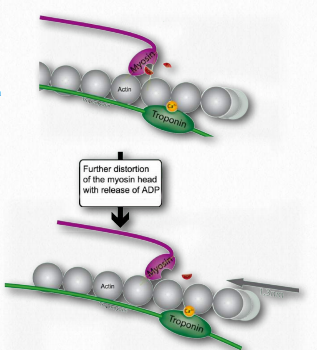
L. Tal movimiento de los filamentos delgados provoca que las líneas Z se acerquen entre sí. Esto da como resultado el acortamiento del sarcómero en su conjunto (Figura 2.122) y el estrechamiento de la banda I y las zonas H (Figura 2.123). M. Si el ATP está disponible, se une a la miosina, lo que le permite soltar la actina (Figuras 2.120 y 2.121). Si el ATP no está disponible, el músculo permanecerá bloqueado en este estado. Esta es la causa del rigor mortis en la muerte - contracción sin liberación de músculos
. 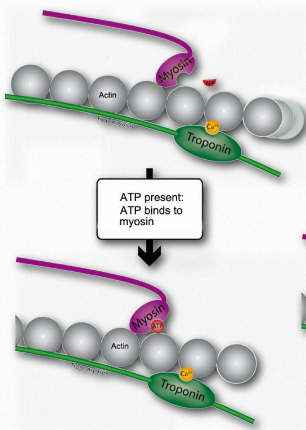
Figura 2.120 - Cuando el ATP está presente, se une a la miosina (M). Wikipedia
N. Después de que la miosina se haya unido al ATP, éste lo hidroliza, produciendo ADP y Pi, los cuales son retenidos por la cabeza. La hidrólisis de ATP restablece la región bisagra a su estado original, flexionándola. Este estado no doblado también se conoce como la posición amarrada.
O.Si la tropomiosina aún permite el acceso a los sitios de unión en la actina, el proceso se repite siempre que el ATP esté disponible y el calcio permanezca a una concentración lo suficientemente alta como para permitir que se una a la troponina.
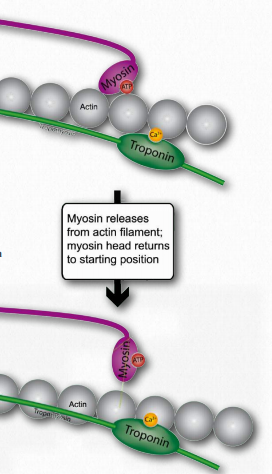
La relajación de la tensión muscular se produce a medida que se disipa el potencial de acción en la célula muscular. Esto sucede porque suceden todas las siguientes cosas 1) la señal nerviosa se detiene; 2) el neurotransmisor es degradado por la enzima acetilcolinesterasa; y 3) la concentración de calcio disminuye porque es absorbido por el retículo sarcoplásmico.
Cabe señalar que el retículo sarcoplásmico siempre está absorbiendo calcio. Solo cuando sus puertas de calcio se abren por el potencial de acción es incapaz de reducir la concentración celular de calcio. A medida que disminuye el potencial de acción, las compuertas de calcio se cierran y el retículo sarcoplásmico “se pone al día” y las concentraciones de calcio celular disminuyen. En ese punto la troponina libera calcio, la tropomiosina vuelve a cubrir los sitios de unión de la miosina en la actina, la miosina pierde su unión a la actina y los filamentos delgados se deslizan de nuevo a sus posiciones originales en relación con los filamentos gruesos de la miosina.
Tropomiosina
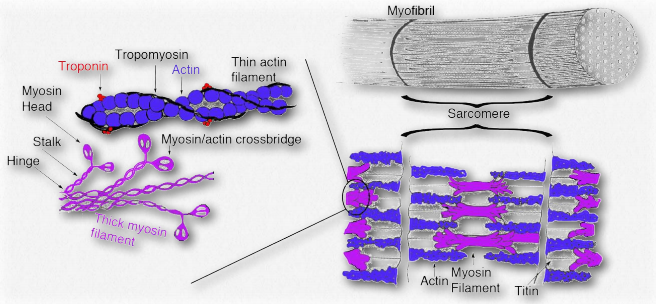
Las tropomiosinas son proteínas que interactúan con filamentos delgados de actina para ayudar a regular sus papeles en el movimiento, tanto en las células musculares como en las células no musculares (Figura 2.124). Las tropomiosinas interactúan para formar dímeros de pies a cabeza y posarse a lo largo de la ranura α-helicoidal de un filamento de actina. Las isoformas de la tropomiosina que se encuentran en las células musculares controlan las interacciones entre la miosina y el filamento de actina dentro del sarcómero y ayudan a regular la contracción del músculo. En otras células, las tropomiosinas no musculares ayudan a regular las funciones del citoesqueleto.
Las interacciones de la tropomiosina con el citoesqueleto son considerablemente más complicadas que lo que ocurre en las células musculares. Las células musculares tienen cinco isoformas de tropomiosina, pero en el citoesqueleto de las células no musculares, hay más de 40 tropomiosinas.
Troponina
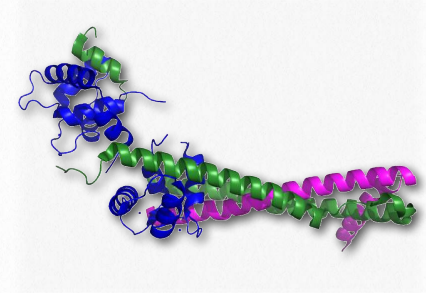
Las troponinas involucradas en la contracción muscular son en realidad un complejo de tres proteínas conocidas como troponina I, troponina C y troponina T (Figura 2.125). Se asocian entre sí y con la tropomiosina en los filamentos de actina para ayudar a regular el proceso de contracción muscular. La troponina I evita la unión de la cabeza de la miosina a la actina y así evita el paso más importante en la contracción.
La troponina C es una unidad que se une a los iones de calcio. La troponina T es responsable de unir las tres proteínas a la tropomiosina. Las troponinas en el torrente sanguíneo son indicativas de trastornos cardíacos. La elevación de las troponinas en la sangre ocurre después de un infarto de miocardio y puede permanecer alta hasta por dos semanas.
Actinina
La actinina es una proteína del músculo esquelético que une filamentos de actina a las líneas Z de las células del músculo esquelético. En las células del músculo liso, también conecta la actina con cuerpos densos.
Titín
La titina (también conocida como conectina) es el equivalente molecular de un resorte que proporciona elasticidad a las células musculares estriadas. Es la tercera proteína más abundante en las células musculares. La proteína es enorme, con 244 dominios proteicos individuales plegados repartidos en 363 exones (mayor número conocido), con el exón más grande conocido (17,106 pares de bases de largo), y es la proteína más grande conocida (27,000 a 33,000 aminoácidos, dependiendo del corte y empalme).
Secuencias no estructuradas
Los dominios proteicos plegados están unidos entre sí por secuencias no estructuradas. Las regiones no estructuradas de la proteína permiten el despliegue cuando se produce el estiramiento y el replegamiento después de la relajación. Titín conecta las líneas M y Z en el sarcómero (Figura 2.123). La tensión creada en la titina sirve para limitar el rango de movimiento del sarcómero, dando lugar a lo que se denomina rigidez pasiva.
Los músculos esqueléticos y cardíacos presentan ligeras variaciones en la secuencia de aminoácidos en sus proteínas de ti estaño y éstas parecen relacionarse con diferencias en las características mecánicas de cada músculo.
Respaldo energético para energía muscular
La mioglobina fue descrita como una masa molecular para oxígeno. Las células musculares tienen una mejor de las suyas para el ATP. El es importante para los animales, pero no para las plantas porque la necesidad de energía de una planta es diferente a la de un animal, las plantas no necesitan acceder a las fuentes de energía tan rápido como los animales, ni tienen que mantener una temperatura interna constante. Las plantas no pueden huir de los depredadores, ni perseguir presas. Estas necesidades de los animales son mucho más inmediatas y requieren que las reservas de energía sean accesibles bajo demanda. Los músculos, por supuesto, permiten el movimiento de los animales y la energía requerida para la contracción muscular es el ATP. Para tener reservas de energía fácilmente disponibles, los músculos tienen, además de ATP, fosfato de creatina para energía y glucógeno para liberación rápida de glucosa para generar más energía. La síntesis de fosfato de creatina es un excelente ejemplo de los efectos de la concentración en la síntesis de moléculas de alta energía. Por ejemplo, el fosfato de creatina tiene una energía de hidrólisis de -43.1 kJ/mol mientras que ATP tiene una energía de hidrólisis de -30.5 kJ/mol. El fosfato de creatina, sin embargo, se elabora a partir de creatina y ATP en la reacción que se muestra en la Figura 2.126. ¿Cómo es esto posible?
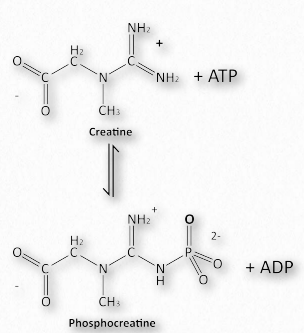
El ∆G°' de esta reacción es +12.6 kJ/mol, reflejando las energías señaladas anteriormente. En una célula muscular en reposo, el ATP es abundante y el ADP es bajo, impulsando la reacción a la baja, creando fosfato de creatina. Cuando comienza la contracción muscular, los niveles de ATP caen y los niveles de ADP suben. La reacción anterior entonces se invierte y procede a sintetizar ATP inmediatamente. Así, el fosfato de creatina actúa como una batería, almacenando energía cuando los niveles de ATP son altos y liberándola casi instantáneamente para crear ATP cuando sus niveles caen.