
10.2: Hacer metazoos
- Page ID
- 53332

A medida que pasamos de la detección de quórum entre organismos del mismo tipo, llegamos a lo que se conoce como biopelículas. Estas son comunidades microbianas, como la placa que se forma en los dientes, y consisten en una serie de diferentes tipos de organismos a menudo codependientes. Si bien la transferencia horizontal de genes puede ocurrir entre estos diferentes organismos, estos permanecen distintos y dan lugar a organismos genéticamente relacionados con su (s) progenitor (s).
El siguiente nivel obvio de organización es lo que llamaremos un organismo colonial discreto. En tales organismos las células individuales se unen entre sí, generalmente a través de los materiales extracelulares que secretan. Obtienen ventajas asociadas con un mayor tamaño (por ejemplo, pueden ser capaces de nadar más rápido o llegar a ser demasiado grandes para tragar fácilmente) pero estas ventajas están restringidas por el hecho de que las células individuales conservan su individualidad.Además, en un organismo colonial puro, cada célula dentro de la colonia conserva su capacidad para reproducirse de forma independiente, ya sea sexual o asexualmente. Anteriormente introdujimos los términos soma para las células del cuerpo que se reproducen asexualmente y son responsables del crecimiento y reparación del organismo, y de la línea germinal, las células que se encargan de producir la próxima generación de organismos. En un organismo puramente colonial, todas las células son células germinales potenciales. No existe un sistema central para coordinar el comportamiento.
Entonces podríamos preguntarnos, ¿cuál es el siguiente paso en la evolución de un organismo multicelular verdaderamente integrado, uno con un soma y una línea germinal? Parece que ha habido una serie de caminos hacia la multicelularidad: por ejemplo, los animales, las plantas y los hongos parecen tener estrategias independientes (linajes). Podemos utilizar organismos modernos como bestiario moderno para ilustrar estas estrategias 310. Esto no es para afirmar que ninguno representa a antepasados reales, todos son organismos modernos, bien adaptados a su entorno actual y al resultado de su propia historia evolutiva. Nunca menos han tratado diversos aspectos de la coordinación y diferenciación multicelular de maneras interesantes.
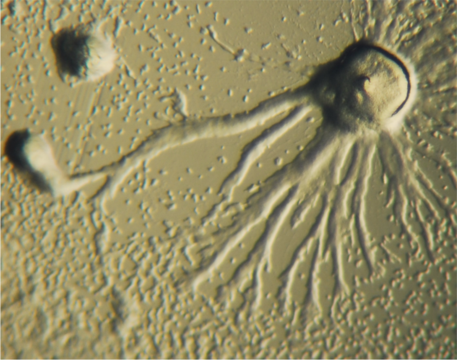
Figura\(\PageIndex{1}\): Dictyostelio que muestra el comportamiento de agregación. Imagen de Wikimedia Commons (“Dictyostelium Aggregation” de Bruno en Columbus — Obra propia. Bajo licencia de dominio público a través de Wikimedia Commons — http://commons.wikimedia.org/ wiki/Archivo:Dictyostelium_agregación.jpg)
Considere el moho de limo eucariota Dictyostelium discoideum. Los mohos celulares de limo viven en el suelo y comen bacterias; son depredadores unicelulares. La mayoría de las veces son células haploides pequeñas, parecidas a amebas. Al morir de hambre pueden someterse a un dramático proceso de agregación. La agregación se desencadena por la liberación de las células individuales de pulsos de adenosina monofosfato cíclico (AMPc); un proceso análogo a la detección de quórum en bacterias (ver arriba). El resultado es que las células individuales comienzan a migrar hacia arriba en el gradiente de concentración de AMPc, donde interactúan y se adhieren entre sí. Los grupos de células producen más AMPc, lo que lleva a la formación de agregados celulares conocidos como babosas que contienen entre 10,000 y 100,000 células discretas. Las babosas migran de manera coordinada. Eventualmente una babosa detendrá su migración e iniciará un proceso conocido como diferenciación, un proceso que implica cambios en la expresión génica y el comportamiento celular. Algunas de las células de la babosa se diferencian para formar células madre. El alargamiento coordinado de estas células madre eleva al aire el resto del “cuerpo” de la babosa. Las células no tallo se diferencian para formar esporas, células como las persistentes quiescentes que mencionamos anteriormente. Cuando se liberan al aire, las esporas están ampliamente dispersas y, si aterrizan en un ambiente apropiado, pueden pasar a formar amebas unicelulares.
A estas alturas ya es posible que pueda generar un escenario plausible para explicar exactamente cómo es posible el comportamiento abnegador de las células madre. La respuesta está en el fitness inclusivo. El propósito de la babosa y el tallo son permitir que las células de Dictyostelium escapen de un ambiente hostil y colonicen ambientes nuevos y más hospitalarios. De hecho, en varios casos las esporas llevan consigo bacterias que inoculan sus nuevos ambientes; estas son bacterias que la ameba puede comer. El moho de limo podría considerarse agricultor bacteriano migratorio 311. Dado que las células ameboides individuales de Dictyostelium no pueden migrar lejos, la mayoría de las células en cualquier región en particular, es decir las células que se combinan para formar una babosa, es probable que estén estrechamente relacionadas entre sí, son parte de un clon. El sacrificio de las células del tallo está más que compuesto por la mayor probabilidad de que las células de esporas sobrevivan y produzcan mucha descendencia. Por supuesto que existe el peligro de que algunas células diverjan (a través de la mutación) y engañen al sistema. Es decir, evitarán convertirse en células del tallo. Tal trampa se ha observado en Dictyostelium de tipo salvaje y el engaño es un desafío que enfrentan todos los sistemas multicelulares. Hay una serie de estrategias que se utilizan para suprimir a los tramposos, generalmente son similares a las explotadas en el contexto de la detección de quórum 312.
Un organismo que muestra un tipo distinto de comportamiento de diferenciación es la bacteria Caulobacter crescentus. Bajo ciertas condiciones producirá un tipo distinto de célula, una célula tallo, que se adhiere a una superficie. Se puededividir para producir células nadadoras que puedan migrar y colonizar nuevas superficies. La célula tallo puede continuar produciendo células nadadoras, y las células nadadoras pueden asentarse y transformarse en células madre. C. crecentus ha establecido dos tipos de células diferentes diseñados para explotar dos entornos distintos.
Referencias
310 El bestiario medieval: http://bestiary.ca
311 Moléculas pequeñas median el cultivo bacteriano por amebas sociales. http://www.ncbi.nlm.nih.gov/pubmed/23975931
El reconocimiento de 312 Kin protege a los cooperadores contra los tramposos: http://www.ncbi.nlm.nih.gov/pubmed/23910661