
6.6: El ciclo de Krebs/TCA/ácido cítrico
- Page ID
- 53788

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)La glucólisis a través de reacciones fermentativas produce ATP anaeróbicamente. La evolución de la respiración (el uso aeróbico de oxígeno para quemar eficientemente los combustibles nutritivos) tuvo que esperar hasta que la fotosíntesis creara la atmósfera oxigénica en la que vivimos ahora. Lee más sobre la fuente de nuestra atmósfera oxigénica en Dismukes GC et al. [(2001) El origen del oxígeno atmosférico en la tierra: la innovación de la fotosíntesis oxigénica. Proc. Nat. Acad. Sci. USA 98:2170-2175].
El ciclo de Krebs es la primera vía de respiración oxígena. La evolución de esta respiración y el puente químico desde la glucólisis hasta el ciclo de Krebs, sin duda ocurrieron algunas reacciones a la vez, quizás al principio como medio para proteger a las células anaeróbicas de los efectos 'venenosos' del oxígeno. Posteriormente, la selección natural desarrolló el ciclo aeróbico de Krebs, el transporte de electrones y las vías de fosforilación oxidativa que vemos hoy en día.
Cualquiera que sea su utilidad inicial, estas reacciones fueron una respuesta adaptativa al aumento de oxígeno en la atmósfera terrestre. Como vía para obtener energía de los nutrientes, la respiración es mucho más eficiente que la glucólisis. Los animales confían en él, ¡pero incluso las plantas y las algas fotosintéticas usan la vía respiratoria cuando la luz solar no está disponible! Aquí nos enfocamos en las reacciones oxidativas en las mitocondrias, comenzando con la oxidación del piruvato y continuando con las reacciones redox del ciclo de Krebs.
Después de ingresar a las mitocondrias, la piruvato deshidrogenasa cataliza la oxidación del piruvato a Acetil-S-Coenzima A (Ac-S-CoA). Entonces el ciclo de Krebs oxida completamente el Ac-S- CoA. Estas reacciones redox mitocondriales generan CO2 y gran cantidad de portadores de electrones reducidos (NADH, FADH2). La energía libre liberada en estas reacciones redox se acopla a la síntesis de un solo ATP por piruvato oxidado (es decir, ¡dos por la glucosa con la que comenzamos!). Es que las moléculas NADH y FADH2 han capturado la mayor parte de la energía libre en las moléculas de glucosa originales. Estas entradas de piruvato en la mitocondria y su oxidación se resumen a continuación.
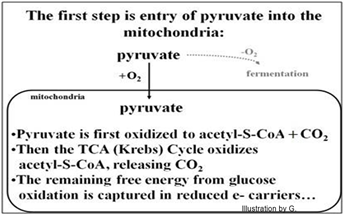
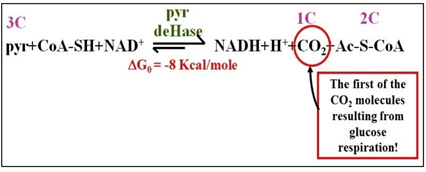
La oxidación del piruvato convierte un carbohidrato 3C en acetato, una molécula 2C, liberando una molécula de CO2. En esta reacción altamente exergónica, CoA-SH forma un enlace tioéster de alta energía con el acetato en Ac-S-CoA. La oxidación del ácido pirúvico resulta en la reducción de NAD+, producción de Ac-S-CoA y una molécula de CO2, como se muestra a continuación.
El ciclo de Krebs funciona durante la respiración para oxidar AC-S-CoA y reducir NAD+ y FAD a NADH y FADH2 (respectivamente). Los intermedios del ciclo de Krebs también funcionan en el metabolismo de aminoácidos y las interconversiones. Todos los organismos aeróbicos vivos hoy en día comparten el ciclo de Krebs que vemos en los humanos. Esto es consistente con su propagación temprana en la evolución de nuestro ambiente de oxígeno. Debido al papel central de los intermedios del ciclo de Krebs en otras vías bioquímicas, algunas partes de la vía pueden incluso haber fechado la vía respiratoria completa. El ciclo de Krebs se desarrolla en mitocondrias de células eucariotas.
Después de la oxidación del piruvato, el Ac-S-CoA ingresa al ciclo de Krebs, condensándose con oxaloacetato en el ciclo para formar citrato. Hay cuatro reacciones redox en el ciclo de Krebs. Al discutir el ciclo de Krebs, busque la acumulación de portadores de electrones reducidos (FADH2, NADH) y una pequeña cantidad de síntesis de ATP por fosforilación a nivel de sustrato. También, siga los carbonos en piruvato en CO2. El Ciclo de Krebs tal como ocurre en los animales se resume a continuación.
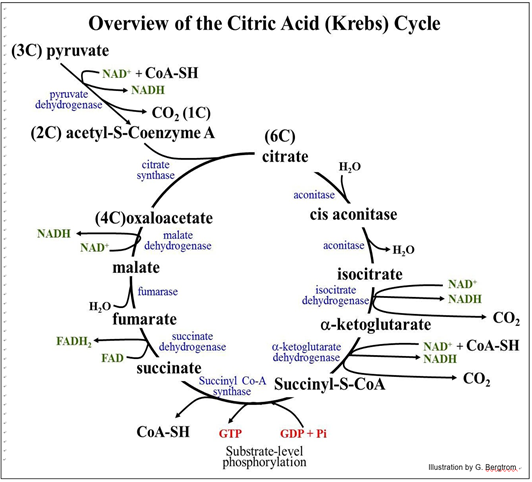
Para ayudarte a entender los eventos del ciclo,
1. encontrar las dos moléculas de CO2 producidas en el propio ciclo de Krebs.
2. encontrar GTP (que transfiere rápidamente su fosfato a ADP para hacer ATP). Tenga en cuenta que en bacterias, el ATP se elabora directamente en este paso.
3. contar todos los portadores de electrones reducidos (NADH, FADH2). Ambos portadores de electrones llevan un par de electrones. Si incluyes los electrones en cada una de las moléculas del NADH hechas en la glucólisis, ¿cuántos electrones se han eliminado de la glucosa durante su completa oxidación?
Recuerde que la glucólisis produce dos piruvatos por glucosa, y así dos moléculas de Ac-S-CoA. Así, el ciclo de Krebs gira dos veces por cada glucosa que ingresa a la vía glucolítica. Los enlaces tioéster de alta energía formados en el ciclo de Krebs alimentan la síntesis de ATP así como la condensación de oxaloacetato y acetato para formar citrato en la primera reacción. Cada NADH lleva alrededor de 50 Kcal de las 687 Kcal de energía libre originalmente disponible en un mol de glucosa; cada FADH2 lleva alrededor de 45 Kcal de esta energía libre. Esta energía alimentará la producción de ATP durante el transporte de electrones y la fosforilación oxidativa.
159 Aspectos destacados del ciclo de Krebs
Por último, ¡la historia del descubrimiento del ciclo de Krebs es tan interesante como el ciclo mismo! Albert Szent-Györgyi ganó un Premio Nobel en 1937 por descubrir algunas reacciones de oxidación de ácidos orgánicos que inicialmente se pensaba que formaban parte de una vía lineal. Hans Krebs realizó los elegantes experimentos demostrando que las reacciones formaban parte de una vía cíclica. Él propuso (¡correctamente!) que el ciclo sería un supercatalizador que catalizaría la oxidación de otro ácido orgánico más. Algunos de los experimentos son descritos por Krebs y sus compañeros de trabajo en su artículo clásico: Krebs HA, et al. [(1938) La formación de ácidos cítrico y α-cetoglutárico en el cuerpo de los mamíferos. Bioquímica. J. 32:113—117]. Hans Krebs y Fritz Lipmann compartieron el Premio Nobel de Fisiología o Medicina 1953. Krebs fue reconocido por su elucidación del ciclo TCA, que ahora lleva más comúnmente su nombre. Lipmann fue reconocido por proponer el ATP como mediador entre la energía alimentaria (nutritiva) y la energía de trabajo intracelular, y por descubrir las reacciones que oxidan el piruvato y sintetizan Ac-S-CoA, uniendo el Ciclo de Krebs y la fosforilación oxidativa (a considerar en el siguiente capítulo).
160 Descubrimiento del ciclo de Krebs
Puedes leer la reseña de Krebs sobre su propia investigación en Krebs HA [(1970) La historia del ciclo del ácido tricarboxílico. Perspectiva. Biol. Med. 14:154-170]. Para leer un clásico sobre cómo Krebs describió su sugerencia de supercatalizador, haga clic en Hans Krebs Comentarios autobiográficos. Para más información sobre la vida de Lipmann, echa un vistazo a la breve nota del Nobel sobre la biografía de Fritz Lipmann.