
14.3: Los 'genes saltares' del maíz
- Page ID
- 54303

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)El informe de Barbara McClintock de que fragmentos de ADN podrían saltar e integrarse en nuevos loci en el ADN fue tan dramático y arcano que muchos pensaron que el fenómeno era único, ¡o no real! ¡Solo con el posterior descubrimiento de transposones en bacterias (y en otros eucariotas) fueron finalmente reconocidos los genes saltarines de McClintock por lo que eran! Al describir sus experimentos, tenga en cuenta que la investigación y las intuiciones de McClintock sobre la regulación génica y la herencia epigenética llegaron mucho antes de que las tecnologías moleculares hicieran posible probar y dar nombres a estos fenómenos. Para comenzar nuestra historia de transposones, mira la ilustración de la reproducción del maíz a continuación.

Los diferentes colores de las semillas de maíz (ker nels) son el resultado de pigmentos de antocianina que son expresados diferencialmente por las células del tejido de aleurona. Mclintock estaba estudiando la herencia de la variación de color, la cual varió de incoloro (blanco o amarillo por ausencia de antocianinas) a marrón, púrpura, manchado o rayado.
El mosaico de colores del grano se muestran vívidamente las mazorcas de maíz en la fotografía de abajo

Claramente, el color del kernel es heredado. La herencia del color de semilla incolora y morada sí siguió las reglas mendelianas, pero la genética del mosaicismo no. Los patrones de color en mosaico después de los cruzamientos genéticos no fueron consistentes, lo que implica que las mutaciones responsables del color del grano no se debieron a mutaciones en células germinales. Más bien, los genes que controlan la síntesis de antocianinas deben estar experimentando mutaciones en células somáticas que se convertirían (o ya eran) en las que se producían los pigmentos.
242 Lo que le interesó a McClintock sobre el maíz
A. Descubrir los genes del mosaicismo; el gen Ds inestable
McClintock buscaba una explicación genética para la variación del color de las semillas en la década de 1940 y principios de 1950. La estructura del ADN solo se había publicado recientemente. ¡La clonación de genes y la secuenciación del ADN estaban décadas en el futuro! Sus únicas tecnologías disponibles se basaron en comprender el surtido alélico mendeliano en estudios de mejoramiento tradicional. Sin embargo, dado que el color de la semilla se expresa en células derivadas del endospermo, ella sabía que la herencia del fenotipo de color del grano debe estudiarse contra un fondo genético triploide. McClintock también estaba al tanto de las propuestas que el fenotipo de color abigarrado podría resultar cuando una 'mutación inestable' que producía granos incoloros 'revertido' en algunas células pero no en otras para crear un fenotipo manchado o rayado. Justo lo que hizo para una 'mutación inestable' era por supuesto, desconocido. McClintock finalmente identificó tres genes involucrados en la coloración del grano de la semilla. Dos de los genes inicialmente estudiados por McClintock controlaron la presencia vs. ausencia de color del grano. Estos son los genes C y Bz:
- C' es el alelo inhibidor dominante, así llamado porque si incluso una copia estaba presente, los granos eran incoloros (amarillos), independientemente del resto del fondo genético
- Bz y bz son alelos dominantes y recesivos del gen Bz, respectivamente. En ausencia de un alelo C' (dominante), la presencia de un alelo Bz conduciría a granos morados. Si el alelo bz estuviera presente sin los alelos tanto C' como Bz, los granos serían de color marrón oscuro.
- El gen requerido para obtener el color abigarrado del grano fue el gen Ds (Disociator). Sin un gen Ds viable, los granos fueron coloreados o incoloros dependiendo de los posibles genotipos dictados por los alelos C y Bz.
En otras palabras, debe ser el gen Ds el que sufre 'mutaciones inestables. Debido a que el efecto del gen Ds fue aleatorio y solo afectó a algunas células de la capa de aleurona, se sospechó que era una región de inestabilidad cromosómica (propensa a daños o roturas) en algunas células pero no en otras. Veamos qué hizo McClintock para averiguar qué estaba pasando en la genética del color del grano de maíz.
Habiendo demostrado ya el cruce en el maíz (en realidad, ¡otro logro notable!) , McClintock mapeó los genes C', Bz y Ds al Cromosoma 9. Luego apareó selectivamente el maíz con los genotipos que se muestran en el siguiente protocolo.
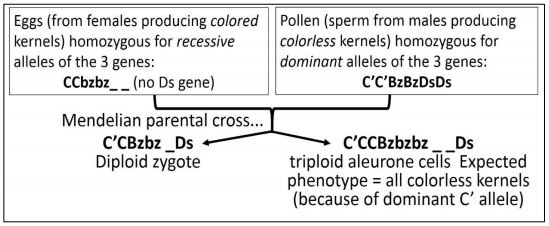
¡Recuerda que los genotipos de células triploides están siendo considerados en esta ilustración! Puede referirse a los efectos fenotípicos de los fondos alélicos de tres genes a medida que seguimos el cruce de McClintock. ¡Su cruz de una recesiva homocigótica con una planta dominante homocigótica debería sonar una campana! Miremos más de cerca esta cruz.
A continuación se muestran los genotipos triploides esperados del cruce.
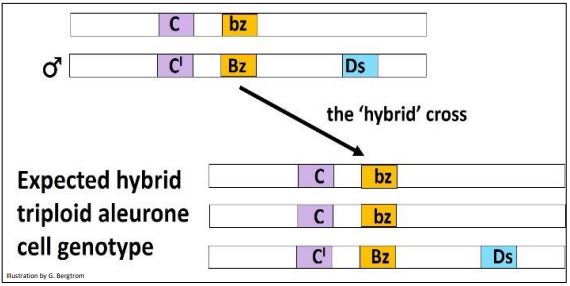
Las células aleuronas resultantes de este cruce deben ser incoloras (amarillas) debido a la presencia del alelo C' dominante. Sin embargo, si bien efectivamente había muchos granos incoloros en la mazorca híbrida, también había muchos granos de mosaico con manchas oscuras/rayas contra un fondo incoloro. La interpretación de los eventos de McClintock se ilustra a continuación
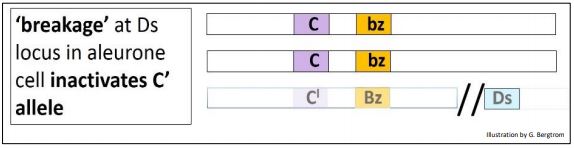
Según McClintock, si algunas células de la capa de aleurona en algunos granos sufrieron rotura cromosómica en el locus Ds (disociador) (indicado por la doble barra,//), el alelo C' se inactiva. Sin un alelo C' funcional, el genotipo operativo en las células afectadas es CCBZbz. Estas células luego vuelven a hacer el pigmento de la ceja como lo indica el alelo bz. Cuando estas células se dividen, crean racimos de células marrones rodeadas de células con un cromosoma ininterrumpido y por lo tanto un alelo C' activo, creando la aparición de manchas o rayas pigmentarias en el núcleo, contra el fondo que de otro modo sería incoloro en las células circundantes.
243 Granos de Maíz Variegados son el resultado de la “Pérdida” del Gen Ds
B. El descubrimiento de genes móviles: el sistema Ac/Ds
Los experimentos que se acaban de describir fueron reproducibles utilizando un único stock reproductivo de maíz. Pero cuando McClintock intentó repetir los experimentos cruzando los machos homocigotos dominantes con hembras homocigóticas recesivas de un criadero diferente, todos los granos de las mazorcas de la progenie eran incoloros, como si el gen Ds no hubiera causado ningún daño cromosómico.
Parecía que el gen Ds aportado por el macho era incapaz de funcionar (es decir, 'romperse') en hembras de este nuevo stock reproductor. McClintock planteó la hipótesis de que la hembra en el cruce original debió haber aportado un factor que de alguna manera podría activar el gen Ds para romperse, y que este factor, otro gen más, estaba ausente o inactivo en las hembras del nuevo criadero. McClintock llamó al nuevo factor el activador, o gen Ac. Con base en la dependencia de Ds en el locus Ac, McClintock reconoció que estos 'genes' formaban parte de un sistema Ac/Ds de 2 elementos que influía en el mosaicismo en granos de maíz.
Luego demostró que la 'rotura' de Ds dependiente de AC se asoció en algunos casos también con la inactivación de un gen Bz normal, lo que condujo a una pérdida de granos de color púrpura. Fue en este punto que McClintock concluyó que lejos de simplemente 'romper' el cromosoma en un locus Ds frágil, el gen Ds en realidad se había movido hacia (o hacia) el gen Bz, interrumpiendo su función. Nuevamente, esto no podría suceder en ausencia de un gen Ac activo. ¡McClintock había descubierto el primer transposón, ganando el Premio Nobel de Fisiología o Medicina de 1983! Con el advenimiento de las tecnologías de ADN recombinante, ahora sabemos que:
- El elemento Ds es un transposón que falta un gen para una enzima transposasa requerida para la transposición.
- El elemento Ac tiene este gen y es capaz de transposición independiente.
- Ac proporciona la transposasa necesaria para movilizarse a sí misma y al elemento Ds
- La similitud de secuencia de los elementos Ds y Ac sustenta su ancestro común.
Las características básicas del sistema Ac/Ds de maíz son:
- Ac tiene 4563 pb de largo.
- Ds es una versión truncada de Ac.
- Hay once pb de repeticiones invertidas en cada extremo del elemento Ac y Ds.
- Hay repeticiones directas de ocho pb (NO repeticiones invertidas) de 'ADN diana' en el sitio de inserción de cualquiera de los transposones.
Busque estas características a medida que describimos los transposones procariotas y eucariotas.