
16.2: Estructura de membrana plasmática
- Page ID
- 54603

En las células eucariotas, la membrana plasmática rodea un citoplasma lleno de ribosomas y orgánulos. Los orgánulos son estructuras que a su vez están encerrados en membranas. Algunos orgánulos (núcleos, mitocondrias, cloroplastos) incluso están rodeados de membranas dobles. Todas las membranas celulares están compuestas por dos capas de fosfolípidos incrustados con proteínas. Todos son selectivamente permeables (semipermeables), permitiendo que solo ciertas sustancias crucen la membrana. Las funciones únicas de las membranas celulares se deben a sus diferentes composiciones de fosfolípidos y proteínas. Décadas de investigación han revelado estas funciones (ver discusiones anteriores sobre la función mitocondrial y cloroplasto, por ejemplo). Aquí describiremos las características generales de las membranas, utilizando la membrana plasmática como nuestro ejemplo.
A. La bicapa fosfolipídica
Gorter y Grendel predijeron la estructura de la membrana bicapa ya en 1925. Sabían que los glóbulos rojos (eritrocitos) no tienen núcleo ni otros orgánulos, y así solo tienen una membrana plasmática. También sabían que el principal componente químico de estas membranas eran los fosfolípidos. El modelo molecular de relleno de espacio a continuación muestra la estructura básica de los fosfolípidos, destacando sus cabezas hidrofílicas (polares) y colas hidrofóbicas.
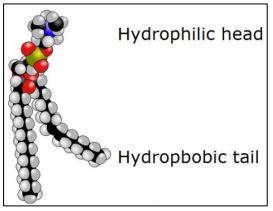
Las moléculas con dominios hidrófilos e hidrófobos son moléculas anfipáticas. Gorter y Grendel habían medido la superficie de los glóbulos rojos. Después hicieron un 'conteo sanguíneo' y luego interrumpieron un número conocido de glóbulos rojos. Luego midieron la cantidad de fosfolípidos en los extractos de membrana. A partir de esto, calcularon que había suficientes moléculas lipídicas por célula para envolverse alrededor de cada célula dos veces. A partir de estas observaciones, predijeron la bicapa fosfolipídica con ácidos grasos interactuando dentro de la bicapa. Curiosamente, Gorter y Grendel habían realizado dos errores de cálculo en la determinación de la cantidad de fosfolípido por célula. Sin embargo, sus errores se compensaban entre sí para que, aunque no estrictamente hablando correcto, ¡su conclusión seguía siendo profética! A continuación se muestran fosfolípidos de membrana comunes.
Las moléculas anfipáticas mezcladas con agua se agregan espontáneamente para 'ocultar' sus regiones hidrofóbicas del agua. ¡En el agua, estos formaron estructuras reales llamadas liposomas que sedimentan cuando se centrifugan!
276 Lípidos de membrana y la bicapa fosfolipídica
277 Experimentos y Usos de Liposomas
La estructura de la membrana liposómica es consistente con la bicapa fosfolipídica predicha, con las colas hidrofóbicas interactuando entre sí y las cabezas polares enfrentadas entre sí, formando una bicapa fosfolipídica. Esto condujo a una imagen de la arquitectura de membrana basada en interacciones de fosfolípidos. A continuación se dibuja una ilustración icónica de la bicapa fosfolipídica, con su interior de ácidos grasos hidrófobos y superficies externas hidrófilas.
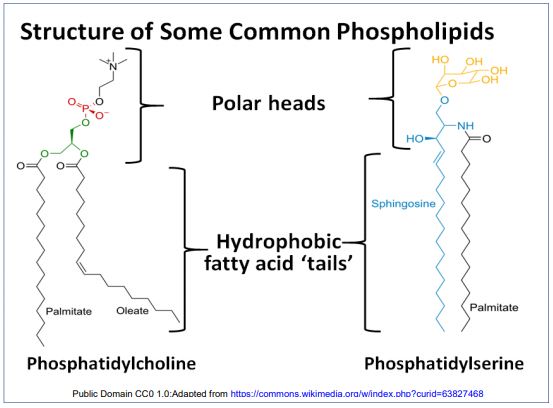
La estructura de la membrana liposómica es consistente con la bicapa fosfolipídica predicha, con las colas hidrofóbicas interactuando entre sí y las cabezas polares enfrentadas entre sí, formando una bicapa fosfolipídica. Esto condujo a una imagen de la arquitectura de membrana basada en interacciones de fosfolípidos. A continuación se dibuja una ilustración icónica de la bicapa fosfolipídica, con su interior de ácidos grasos hidrófobos y superficies externas hidrófilas.
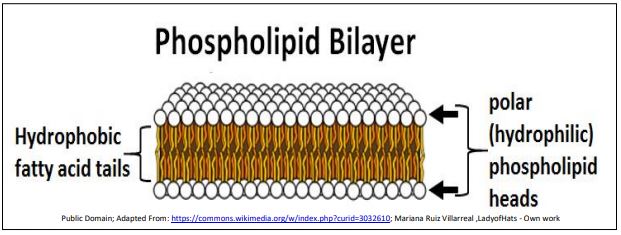
B. Modelos de Estructura de Membrana
En 1935, Davson y Danielli sugirieron que las proteínas podrían unirse a las cabezas polares de los fosfolípidos en la membrana plasmática, creando un sándwich proteína/lípido/proteína. Décadas después, J.D. Robertson observó membranas en el microscopio electrónico de transmisión a alta potencia, revelando que todas las membranas celulares tenían una estructura trilamelar. A continuación se ilustra el aspecto trilamelar clásico de una membrana celular en el microscopio electrónico
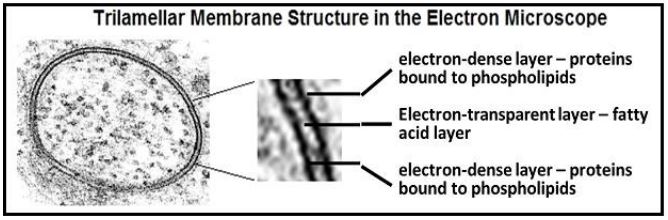
La estructura trilamelar es consistente con las superficies hidrófilas recubiertas de proteína de una bicapa de fosfolípidos en el sándwich proteína-lípido-proteína de Davson y Danielli. Al observar que todas las membranas celulares tenían esta estructura trilamelar, Robertson propuso además su modelo Unit Membrane: todas las membranas consisten en una bicapa de fosfolípidos transparente recubierta con proteínas densas en electrones.
La vista estática de los modelos trilamelares de estructura de membrana implícita en los modelos Davson-Danielli o Robertson fue reemplazada en 1972 por el modelo Fluid Mosaic de Singer y Nicolson (ver El modelo de mosaico de fluidos de membranas. Ciencia 175:720 - 731). Sugirieron que además de las proteínas periféricas que sí se unen a las superficies de las membranas, muchas proteínas integrales de membrana realmente abarcan la membrana. Las proteínas integrales de membrana se imaginaron como un mosaico de 'azulejos' proteicos incrustados en un medio fosfolípido. Pero a diferencia de un mosaico de azulejos esmaltados en una estructura firme, similar al cemento, se predijo que las 'tejas' de proteínas serían móviles (fluidas) en un mar de fosfolípidos. En este modelo, las proteínas de membrana están ancladas en membranas por uno o más dominios hidrófobos; sus dominios hidrófilos enfrentarían ambientes acuosos externos y citosólicos. Así, al igual que los propios fosfolípidos, las proteínas de membrana son anfipáticas. Sabemos que las células exponen diferentes características estructurales (y funcionales) superficiales al ambiente acuoso en lados opuestos de una membrana. Por lo tanto, también decimos que las membranas celulares son asimétricas. A continuación se ilustra un modelo típico de la membrana plasmática de una célula.
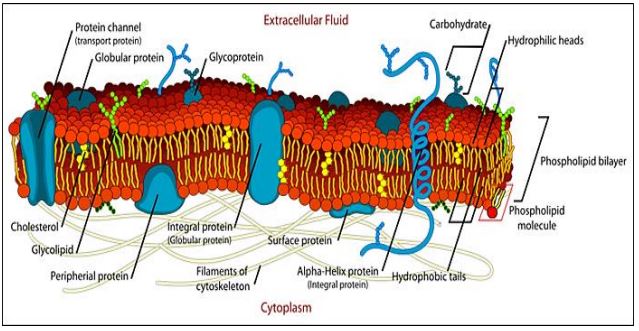
En este modelo, las proteínas periféricas tienen un dominio hidrófobo que no abarca la membrana, sino que la ancla a un lado de la membrana. Otras proteínas periféricas (o llamadas “de superficie”) se unen a la membrana mediante interacciones con los grupos fosfato polares de los fosfolípidos, o con los dominios polares de las proteínas integrales de membrana.
Debido a sus propios dominios hidrofílicos acuosos, las proteínas de membrana son una barrera natural para el paso libre de moléculas cargadas a través de la membrana. Por otro lado, las proteínas de membrana son responsables de la permeabilidad selectiva de las membranas, facilitando el movimiento de moléculas específicas dentro y fuera de las células. Las proteínas de membrana también explican interacciones específicas y selectivas con su entorno extracelular. Estas interacciones incluyen la adhesión de las células entre sí, su unión a las superficies, la comunicación entre las células (tanto directa como a través de hormonas y neuronas), etc. El 'recubrimiento de azúcar' de las superficies extracelulares de las membranas plasmáticas proviene de oligosacáridos unidos covalentemente a la membrana proteínas (como glicoproteínas) o a fosfolípidos (como glicolípidos). Los componentes carbohidrato de las proteínas de membrana glicosiladas informan su función. Así, las glicoproteínas permiten interacciones específicas de las células entre sí para formar tejidos. También permiten la interacción con superficies extracelulares a las que deben adherirse. Además, ocupan un lugar destacado como parte de los receptores de muchas hormonas y otras biomoléculas de comunicación química. Los dominios proteicos expuestos al citoplasma, aunque no están glicosilados, a menudo se articulan a componentes del citoesqueleto, dando a las células su forma y permitiendo que las células cambien de forma cuando sea necesario. Muchas proteínas de membrana tienen características enzimáticas esenciales, como veremos. Dado el papel crucial de las proteínas y glicoproteínas en la función de la membrana, no debería sorprender que las proteínas constituyan un promedio de 40-50% de la masa de una membrana. En algunos casos, las proteínas representan hasta el 70% de la masa de la membrana (¡piense en las membranas de cristal en las mitocondrias!).
278 Propiedades de las proteínas incrustadas en una bicapa fosfolipídica
C. Evidencia de la estructura de membrana
La asimetría de membrana se refiere a las diferentes características de la membrana enfrentadas a lados opuestos de la membrana. Esto se demostró directamente mediante la técnica de microscopía electrónica de barrido de congelación-fractura. La técnica consiste en congelar membranas aisladas en agua y luego astillar el hielo. Cuando el hielo se agrieta, las membranas envueltas se parten a lo largo de una línea de menor resistencia... que resulta estar entre las colas opuestas de ácidos grasos hidrófobos en el interior de la membrana. Luego, la microscopía electrónica de barrido revela las características de las superficies de la membrana interior y exterior. Entre las características destacadas en una micrografía de barrido de membranas plasmáticas fracturadas por congelación se encuentran los fosos y montículos opuestos enfrentados entre sí en colgajos opuestos de la membrana, como se ilustra a continuación.
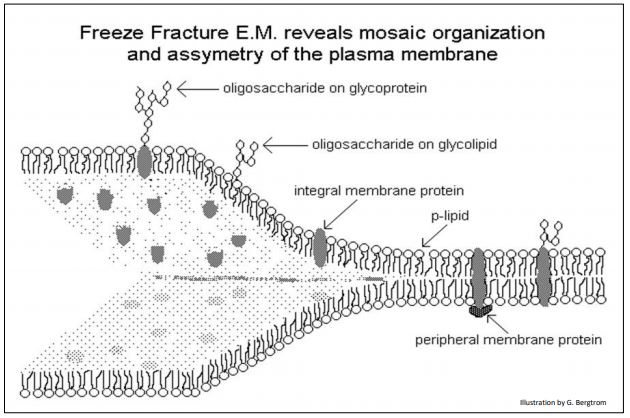
Otras características que se muestran aquí son consistentes con la estructura de membrana de fosfolípidos.
280 Microscopía electrónica por fractura por congelación de membranas celulares
La citoquímica confirmó la asimetría de la membrana plasmática, mostrando que solo las superficies externas de las membranas plasmáticas están recubiertas de azúcar, Consulte el siguiente enlace para obtener descripciones más detalladas de los experimentos.
281 EM Demostración Citoquímica de Asimetría de Membrana
Finalmente, también se demostró bioquímicamente la asimetría de las membranas. En un experimento, las células enteras se trataron con enzimas proteolíticas, seguido de extracción de las membranas y luego aislamiento de proteínas de membrana. En un segundo experimento, primero se aislaron membranas plasmáticas de células no tratadas y luego se trataron con las enzimas. En un tercer experimento, se extrajeron proteínas de membranas plasmáticas aisladas de células no tratadas. La separación electroforética de los tres extractos proteicos por tamaño demostró que diferentes componentes de proteínas integrales de membrana estuvieron presentes en los dos experimentos de digestión, confirmando la asimetría de la membrana plasmática. Nuevamente, para más detalles, consulta el siguiente enlace.
282 Demostración Electroforética de Asimetría de Membrana
También se probó la idea de que las membranas son fluidas. En otro experimento elegante, se fabricaron anticuerpos contra proteínas de membrana celular de ratón y humana. Las membranas se aislaron e inyectaron en un tercer animal (probablemente un conejo). El conejo vio las membranas y sus proteínas asociadas como extrañas y respondió haciendo moléculas específicas de anticuerpos anti-membrana. Los anticuerpos contra cada fuente de membrana se aislaron y se etiquetaron por separado con etiquetas fluorescentes de diferentes colores para que brillaran de un color diferente cuando se sometieran a luz ultravioleta. Después se mezclaron células de ratón y humanas en condiciones que provocaron que se fusionaran, haciendo células híbridas humano-ratón. Cuando se agregan a las células fusionadas, los anticuerpos etiquetados se unen a las proteínas de la superficie celular. Al poco tiempo, se observó que los diferentes anticuerpos fluorescentes se mezclaban bajo un microscopio de fluorescencia bajo luz UV. Las etiquetas fluorescentes parecían moverse de su ubicación original en las membranas fusionadas. Claramente, las proteínas incrustadas en la membrana no son estáticas, sino que son capaces de moverse lateralmente en la membrana, en efecto flotando y difundiendo en un “mar de fosfolípidos”. Los anticuerpos de ratón como se ven en la célula híbrida justo después de la fusión se cartografian a continuación.
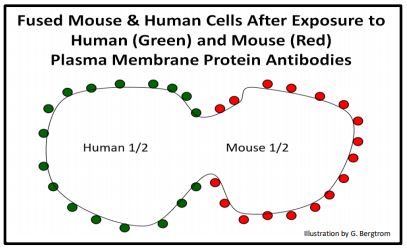
283 Dos demostraciones de fluidez de la membrana: el mosaico de fluidos
D. La fluidez de la membrana está regulada
1. Factores químicos que afectan la fluidez de la membrana
Como se podría imaginar, la fluidez de una membrana depende de su composición química y de las condiciones físicas que rodean a la célula, por ejemplo la temperatura exterior. A continuación se resumen los factores que afectan la fluidez de la membrana.
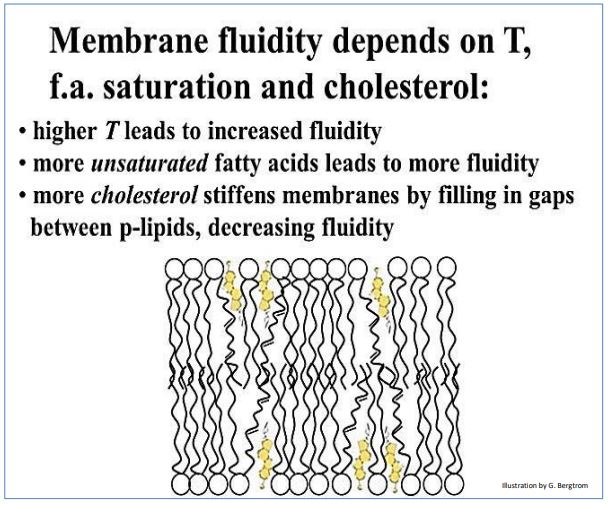
Así como calentar una solución hace que las moléculas disueltas y las partículas se muevan más rápido, los componentes fosfolípidos y proteicos de las membranas también son más fluidos a temperaturas más altas. Si los ácidos grasos de los fosfolípidos tienen más enlaces de carbono insaturados (C=C), estas colas hidrófobas tendrán más torceduras, o curvas. Los torceduras tienden a separar las colas de fosfolípidos. Con más espacio entre las colas de ácidos grasos, los componentes de la membrana pueden moverse más libremente. Así, más ácidos grasos poliinsaturados en una membrana la hacen más fluida. Por otro lado, las moléculas de colesterol tienden a llenar el espacio entre los ácidos grasos en el interior hidrófobo de la membrana. Esto reduce la movilidad lateral de los componentes fosfolípidos y proteicos en la membrana. Al reducir la fluidez, el colesterol reduce la permeabilidad de la membrana a algunos iones.
2. Factores funcionales que afectan la fluidez de la membrana
La evolución ha adaptado las membranas celulares a diferentes y cambiantes ambientes para mantener la fluidez necesaria para el correcto funcionamiento celular. Los organismos poiquilotérmicos o de sangre fría, desde procariotas hasta peces y reptiles, no regulan su temperatura corporal. Así, cuando se exponen a temperaturas más bajas, las poiquilotermas responden aumentando el contenido de ácidos grasos insaturados de sus membranas celulares. A temperaturas más altas, aumentan el contenido de ácidos grasos saturados de membrana. Así, las membranas celulares de peces que viven bajo el hielo ártico mantienen la fluidez al tener altos niveles de ácidos grasos tanto monoinsaturados como poliinsaturados. ¿Qué pasa con las especies de peces que varían en ambientes más cálidos y fríos (o que viven en climas con estaciones cambiantes)? Para estos peces, la composición de la membrana puede cambiar para ajustar la fluidez al ambiente.
Los mamíferos y aves de sangre caliente (homeotérmicos) mantienen una temperatura corporal más o menos constante. Como resultado, su composición de membrana también es relativamente constante. ¡Pero hay una paradoja! Sus membranas celulares son muy fluidas, con una mayor proporción de grasa poliinsaturada a grasas monoinsaturadas que digamos, reptiles. La aparente paradoja se resuelve, sin embargo, cuando entendemos que esta mayor fluidez apoya la mayor tasa metabólica de las especies de sangre caliente en comparación con los poiquilotermos. ¡Solo compara los estilos de vida de casi cualquier mamífero con un cocodrilo flotante perezoso, o una serpiente que disfruta a la sombra de una roca!
E. Elaboración y Experimento con Membranas Artificiales
Las estructuras similares a membranas se pueden formar espontáneamente. Cuando los fosfolípidos interactúan en un ambiente acuoso, se agregan para excluir sus colas grasas hidrófobas del agua, formando micelas. Las micelas son vesículas esféricas monocapa de fosfolípidos que se autoensamblan, una agregación natural de los dominios de ácidos grasos hidrófobos de estas moléculas anfipáticas.
A continuación se dibuja una micela.

Las micelas pueden autoensamblarse adicionalmente en bicapas esféricas de fosfolípidos llamadas liposomas (abajo).

Cuando se forman en el laboratorio, estas estructuras se comportan un poco como células, por ejemplo, formando un pellet en el fondo de un tubo cuando se centrifugan. Los liposomas se pueden diseñar a medida a partir de diferentes tipos de fosfolípidos y proteínas anfipáticas que se vuelven integrales a las membranas de los liposomas. Cuando los liposomas se pueden preparar en presencia de proteínas específicas u otras moléculas que no pueden cruzar la membrana. Las moléculas atrapadas no pueden salir de este 'organelle' sintético. Tales fueron los estudios que permitieron la identificación de los complejos mitocondriales de la cadena respiratoria. La capacidad de manipular el contenido de liposomas y la composición de la membrana también los convierte en candidatos para la administración de fármacos a células y tejidos específicos (google liposomas para más información).
F. La Membrana Plasma está Segragada en Regiones con Diferentes Propiedades de Fluidez y Permeabilidad Selectiva
Como veremos en breve, la fluidez no da como resultado una difusión igual de todos los componentes de la membrana alrededor de la superficie de la membrana celular. En cambio, las conexiones extracelulares entre las células, así como las conexiones intracelulares de la membrana con regiones diferenciadas del citoesqueleto, compartimentan efectivamente la membrana en subregiones. Para entender esto, imagina una lámina de epiteliales como las de la caricatura de abajo.
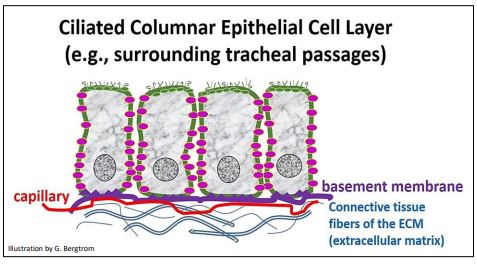
La lámina de células expone una superficie con funciones únicas al interior del órgano que alinean. Expone la superficie opuesta, una con una función bastante diferente, al otro lado de la lámina. Las superficies laterales de las células son otro compartimento de membrana, uno que funciona para conectar y comunicarse entre las células en la lámina. Los componentes, es decir, proteínas de membrana ilustradas con diferentes formas y colores simbólicos, pueden permanecer fluidos dentro de un compartimento. Por supuesto, esta macrodiferenciación de las membranas celulares para permitir interacciones célula-célula y célula-ambiente tiene sentido intuitivo.
La observación reciente de que las membranas celulares están aún más compartimentadas fue quizás menos anticipada. De hecho, las membranas se dividen en microcompartimentos. Dentro de estos compartimentos, los componentes son fluidos pero rara vez se mueven entre compartimentos. Los estudios indican que los elementos citoesqueléticos crean y mantienen estas microdiscontinuidades. Por ejemplo, las proteínas integrales de membrana se inmovilizan en las membranas si están unidas a fibras citoesqueléticas (p. ej., actina) en el citoplasma. Además, cuando los agregados de estas proteínas se alinean debido a interacciones similares, forman una especie de valla, inhibiendo el cruce de otros componentes de la membrana. Por analogía, este mecanismo de microcompartimentalización se llama el modelo de Cercas y Piquetes; las proteínas adheridas al citoesqueleto sirven como piquetes. El movimiento a través de las vallas (es decir, de un compartimento de membrana a otro) es poco frecuente. Presumiblemente se necesita energía cinética adicional para que una molécula 'salte' una valla entre compartimentos. De ahí que este tipo de movimiento, o difusión del lúpulo, lo distingue del movimiento browniano implícito en el modelo original de mosaico fluido.
285 Dominios de membrana: Diferenciación regional de una membrana plasmática