17.2: Transporte por Membrana
- Page ID
- 54459

El primer control sobre el paso de moléculas a través de las membranas es el carácter semipermeable de la membrana misma. Las moléculas se mueven dentro y fuera de las células en una de tres formas: difusión pasiva, transporte facilitado y transporte activo.
Solo unas pocas moléculas pequeñas y relativamente sin carga pueden atravesar una membrana sin ayuda (es decir, por difusión pasiva). Las moléculas hidrofílicas que deben entrar o salir de las células lo hacen con ayuda, es decir, mediante el transporte facilitado. El transporte pasivo y facilitado libera la energía libre inherente a los gradientes de concentración mientras las moléculas se difunden a través de una membrana. En contraste, el transporte activo consume energía para crear gradientes de concentración de solutos específicos. La especificidad del transporte facilitado y activo radica en proteínas integrales de membrana que reconocen y se unen a solutos específicos para el transporte. Como puedes predecir, la regulación alostérica de estas proteínas controla el movimiento de sus moléculas diana dentro o fuera de las células.
A pesar de su polaridad, muchos creían que las pequeñas moléculas de agua cruzaban membranas sin ayuda. En efecto, lo hace de manera limitada. Sin embargo, otros sospechaban que dado sus enlaces covalentes polares altamente cargados en relación con su pequeño tamaño, las moléculas de agua requieren una ayuda para atravesar las membranas de manera eficiente. Comencemos con una mirada más cercana a la difusión pasiva y la difusión facilitada, seguida de la ósmosis (un caso especial de difusión facilitada), y finalmente, al transporte activo.
A. Difusión Pasiva
La difusión pasiva es el movimiento de las moléculas a lo largo del tiempo por movimiento aleatorio (también llamado movimiento browniano) desde regiones de mayor concentración hasta regiones de menor concentración. La difusión pasiva significativa a través de las membranas celulares se limita a unas pocas moléculas, en su mayoría gases como O2, CO2 y N2, que pueden cruzar libremente la barrera hidrofóbica de fosfolípidos. La rápida difusión de gases es esencial para el intercambio de O2 y CO2 entre los capilares alveolares y las células de los pulmones durante la respiración fisiológica. El intercambio de O2 y CO2 también ocurre en las mitocondrias durante la respiración celular. La difusión a través de las membranas no requiere energía. De hecho, la difusión libera energía, recuerda el movimiento de protones a través de la puerta de protones ATPasa F1 que sintetiza ATP durante la fosforilación oxidativa mitocondrial.
La velocidad de difusión de una molécula depende únicamente de su propia concentración. No se ve afectada por la concentración de otras moléculas. Con el tiempo, el movimiento aleatorio de solutos dentro y a través de los compartimentos da como resultado un equilibrio dinámico para cada soluto diferente a lo largo del tiempo. En equilibrio, las moléculas de soluto continúan difundiéndose a través de la membrana, pero para cada molécula que se mueve en una dirección, otra molécula del mismo soluto cruza en la otra dirección.
B. Difusión facilitada de solutos e iones
Al igual que la difusión pasiva, la difusión facilitada es el paso espontáneo (cuesta abajo) de moléculas o iones a través de membranas a través de proteínas transmembrana específicas. La cinética de difusión pasiva y facilitada revela las diferencias entre los dos procesos. Para entender esto último, recordemos que la tasa de catálisis enzimática es saturable. Es decir, a medida que aumenta la concentración de sustrato, la velocidad de la reacción catalizada se aproxima a un máximo (V máx), cuando todas las moléculas enzimáticas en solución se unen a moléculas de sustrato. El mismo fenómeno de saturación se aplica al transporte facilitado: la velocidad de movimiento del soluto a través de una membrana es directamente proporcional al número de proteínas de transporte en la membrana.
La cinética de difusión pasiva y facilitada se ilustra mediante el gráfico que se muestra a continuación
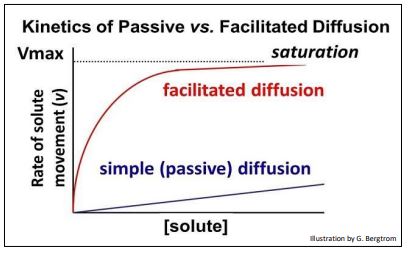
¡Quizás veas otra similitud entre la difusión facilitada y la catálisis enzimática en esta gráfica! Las tasas relativas de difusión facilitada suelen ser rápidas, en comparación con las de difusión pasiva. Esto se debe a que los cambios alostéricos que acompañan al transporte facilitado son rápidos, tal como lo son durante la catálisis enzimática. Hay tres tipos de transport facilitado de solutos (abajo).
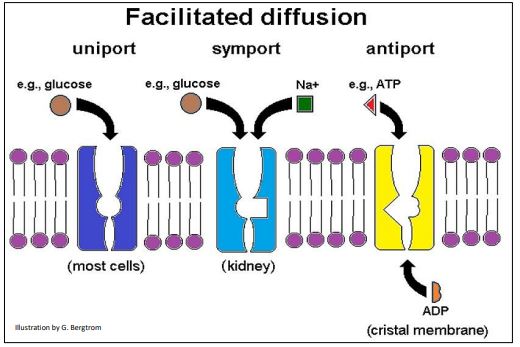
La proteína GLUT (transportador de glucosa) que se muestra arriba (izquierda) permite unipuerto de glucosa, el transporte específico de una sola sustancia dentro o fuera de las células. Las células renales tienen transportadores de glucosa que simportan (acoplan) el movimiento simultáneo de los iones de glucosa y sodio; SGLT (S-odium- GL ucose T ransporter) cumple una función similar en las células del intestino delgado, permitiendo la absorción de glucosa en la dieta y sodio. Antiport (arriba, derecha) permite el intercambio específico de moléculas a través de una membrana. En el ejemplo mostrado, el ATP sale de la matriz mitocondrial, cruzando la membrana cristalina al mismo tiempo que el ADP ingresa a la matriz.
Ya sea por unipuerto, simporte o antipuerto, cada soluto atravesará independientemente una membrana por su gradiente de concentración, moviéndose de mayor concentración a donde se encuentra a una concentración menor. Recordemos que la difusión a lo largo de un gradiente libera energía libre que depende de las concentraciones relativas de los solutos.
Las proteínas que median la difusión facilitada son de dos tipos: proteínas portadoras y proteínas de canal. Las proteínas transportadoras permiten el transporte de solutos. Los iones, con su alta relación de carga a masa, necesitan ayuda para cruzar la barrera de membrana hidrófoba; este es el trabajo de las proteínas de canal que esencialmente sirven como poros iónicos.
Al igual que todas las proteínas transportadoras, tanto las proteínas portadoras como las de canal experimentan cambios alostéricos durante el transporte. También suelen estar sujetos a regulación alostérica, en lugar de estar en un estado constante 'abierto'. Ejemplos de difusión facilitada se consideran con más detalle a continuación.
1. Proteínas Transportadoras
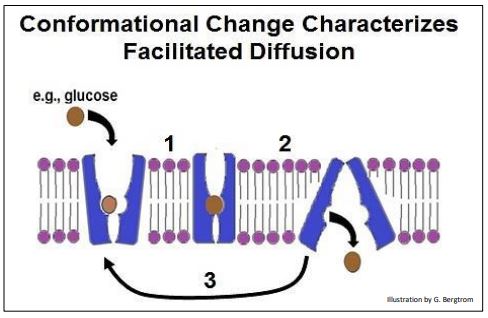
Cuando una proteína portadora se une a un soluto que debe atravesar la membrana, sufre un cambio alostérico (ilustrado a continuación). Durante el transporte, la proteína portadora sufre otro cambio de forma. Cuando el soluto alcanza el otro lado de la membrana, ya no tiene una alta afinidad por la proteína portadora. Después de la liberación del soluto, un cambio alostérico final restaura la conformación original de la proteína de transporte. Estos cambios conformacionales secuenciales se ilustran en la página siguiente.
Cualquier proteína portadora dada es específica para un solo soluto, o como mucho una sola familia de solutos estrechamente relacionados. Acabamos de ver la proteína transportadora GLUT1 que permite la glucosa (¡pero no fructosa ni ribosa!) para cruzar las membranas celulares. Diferentes proteínas portadoras específicas facilitan el transporte de aminoácidos y otros solutos cargados a través de las membranas celulares. Una vez más, las moléculas que indican el estado celular (es decir, la necesidad de importar o exportar soluto) son efectores alostéricos que regulan las proteínas portadoras. La regulación del transporte de glucosa a las células por la insulina es una perfecta
ejemplo. Una consecuencia de la insulina liberada durante una comida (o simplemente antes de una comida) es la estimulación de los transportadores de glucosa para que absorban la glucosa. La incapacidad de esos transportadores para responder a la insulina explica en parte la diabetes tipo II (inicio adulto).
El agua atraviesa las membranas por ósmosis (veremos más de cerca cómo la ósmosis afecta a las células en un momento). Observamos que pequeñas cantidades de agua podrían cruzar la bicapa de fosfolípidos sin ayuda. El agua también puede atravesar una membrana incidentalmente, cuando los iones fluyen a través de sus proteínas de canal. Pero la mayor parte de la ósmosis implica difusión facilitada mediada por acuaporinas. Algunas acuaporinas solo transportan agua. Otros han evolucionado para cofacilitar el transporte de glucosa (ver arriba), glicerol, urea, amoníaco, dióxido de carbono e incluso iones (protones) junto con el agua. Al igual que otras proteínas portadoras, las acuaporinas están reguladas alostéricamente para permitir que las células cumplan con sus requisitos específicos de equilibrio hídrico. Tan fundamental fue la comprensión del balance hídrico que el descubrimiento de las acuaporinas le valió a Peter Agre un Premio Nobel de Química en 2003. Desde el descubrimiento de Agre (en 1992), varias enfermedades genéticas se han relacionado con mutaciones en el gen de la acuaporina.
Las células renales están involucradas críticamente en el equilibrio hídrico de los vertebrados y tienen muchas acuaporinas en sus membranas. En una forma poco frecuente de diabetes, las acuaporinas anormales hacen que los riñones excreten volúmenes de agua inusualmente grandes. En otro ejemplo, las mutaciones del gen de la acuaporina conducen al desarrollo de cataratas en ambos ojos. Desde su descubrimiento inicial, las acuaporinas se han descrito en bacterias y plantas. Para obtener más información, haga clic en Acuaporinas.
2. Canales Iónicos
La regulación alostérica de las proteínas de los canales iónicos controla la homeostasis iónica en sangre y fluidos extracelulares dentro de límites estrechos. A menudo, múltiples proteínas integrales contribuyen a la formación de un canal iónico. Cuando se estimulan, las proteínas del canal se reorganizan para abrir un poro permitiendo el transporte específico de iones. Algunos canales iónicos, como el sistema simport iónico glucosódico señalado anteriormente, movilizan la energía de difusión de un soluto (un ion en este caso) para transportar rápidamente otro soluto a través del mismo canal (actuando como un canal iónico y una proteína portadora). Finalmente, los canales iónicos son responsables de la excitabilidad de las células, donde los canales de Na+, K+ y Ca++ colaboran en movimientos iónicos dentro y fuera de las células que conducen a respuestas celulares neuronales o musculares (¡más en breve!)
C. Ósmosis
La ósmosis, la difusión del agua a través de las membranas de concentraciones de soluto menores a mayores, es una actividad esencial. Permite que las células utilicen agua para mantener la integridad celular o adaptarse a los cambios en la composición del soluto en el ambiente extracelular. La ósmosis a través de las membranas celulares se basa en el transporte facilitado de agua por las acuaporinas. La difusión pasiva de moléculas de agua, se puede demostrar con una membrana artificial (por ejemplo, diálisis). El agua cruzará dicha membrana si las concentraciones de soluto son mayores en un lado de la membrana. El agua cruza la membrana “tratando” de igualar las concentraciones de soluto en ambos lados de la membrana. En efecto, el movimiento del agua es desde el lado de una membrana donde la concentración de moléculas de agua libre es mayor (es decir, donde la concentración de soluto es menor) hasta el lado donde la concentración de agua libre es menor (es decir, donde la concentración de soluto es mayor).
1. Ósmosis en células vegetales y animales
Podríamos presentar esta sección en el contexto de las concentraciones de agua libre, pero lo haremos en términos más familiares de concentraciones de soluto. La ósmosis afecta a las células vegetales y animales de acuerdo con los mismos principios, pero con diferentes efectos. Primero, consideremos el efecto de diferentes concentraciones experimentales de soluto en células animales, ilustradas en la página siguiente.
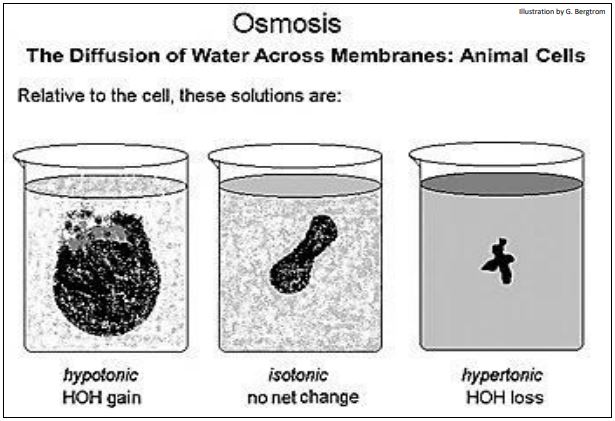
Si la concentración de soluto dentro y fuera de la célula es la misma, no hay movimiento neto dentro o fuera de las células. Se dice que el medio extracelular y el citosol son isotónicos entre sí. Cuando el agua se difunde hacia las células desde un medio bajo en soluto, se dice que el medio es hipotónico a (menos concentrado que) el citosol. En este caso, el movimiento del agua hacia una célula disminuye la concentración de soluto del citosol. Las células animales se hinchan y estallan en una solución hipotónica. En soluciones hipertónicas (con concentraciones de soluto mayores que el citosol), las células animales se marchitan a medida que el agua sale de la célula. A partir de esta breve descripción, se debe concluir que el agua cruza desde el lado hipotónico hasta el lado hipertónico de una membrana.
Al igual que con las células animales, la exposición de las células vegetales a soluciones hipotónicas o hipertónicas provoca los mismos movimientos direccionales del agua, pero con algunas diferencias clave debido a sus paredes celulares. En soluciones hipotónicas, el agua ingresa a las células vegetales, moviéndose hacia el citosol y luego hacia las vacuolas de agua llamadas tonoplastos. Esto da como resultado una mayor presión osmótica (presión de agua) en los tonoplastos. El tonoplasto en expansión crea presión de turgencia, comprimiendo el citosol contra la pared celular. En lugar de estallar, las células y los tejidos vegetales se endurecen y se vuelven turgentes. Dado que el agua no puede ingresar indefinidamente a las células vegetales, el agua deja de ingresar a las células cuando la presión osmótica fuera de las células y la presión de turgencia dentro de las células están en equilibrio. Te encontraste con este fenómeno si alguna vez has regado demasiado las plantas de interior. Las hojas y tallos rígidos se vuelven quebradizos y se rompen o rompen fácilmente. En medio hipertónico, las células vegetales (como las células animales) pierden agua, lo que resulta en plasmólisis. Este es el efecto de la contracción de la membrana plasmática. Sin embargo, la membrana plasmática permanece firmemente unida a la pared celular de la planta en varios puntos. Es posible que hayas visto plantas poco regadas con tallos y hojas flojos o caída. Estos se han vuelto flácidos debido a la pérdida de agua y, por lo tanto, a la pérdida de presión de turgencia necesaria para mantener las hojas y los tallos erguidos. Los efectos de diferentes soluciones sobre las células vegetales se ilustran a continuación.
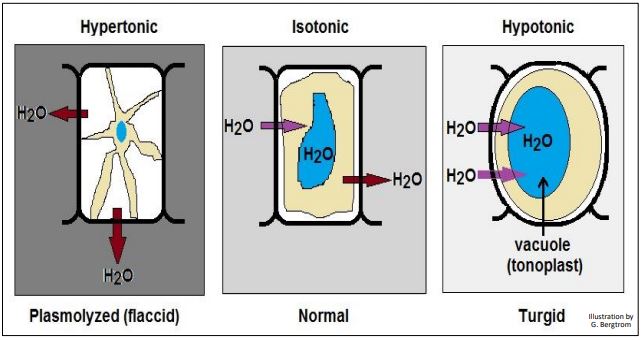
Formalmente, la presión osmótica o turgencia se define como la fuerza por unidad de área (es decir, presión) requerida para evitar el paso del agua a través de una membrana semipermeable de una solución hipotónica a una hipertónica
2. Ósmosis en la vida vegetal
Mientras que las células vegetales individuales responden a cambios en las concentraciones de solutos, estos cambios se comunican rápidamente a las células adyacentes a través de plasmodesmas. Estas estructuras conectan las membranas plasmáticas de las células adyacentes a través de sus paredes celulares, permitiendo un intercambio rápido y directo de información física y química.
A continuación se ilustra un plasmodesma.
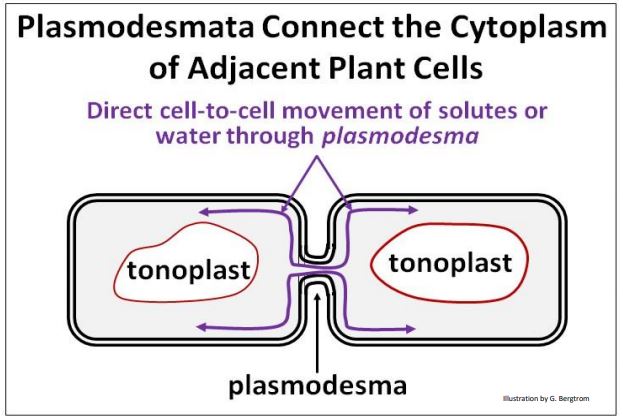
De esta manera, los efectos sobre la presión osmótica en unas pocas células creados por los cambios en la disponibilidad de agua se transmiten a las células adyacentes, afectando la presión de turgencia en esas células y, en última instancia, en los tejidos vegetales.
Por último, ¡la vida vegetal depende del agua! Recordemos que las células vegetales requieren un suministro continuo de agua para su uso en la fotosíntesis, para proporcionar hidrógeno para reducir el CO2 a la glucosa. La fotosíntesis así como la pérdida de exceso de agua de los tejidos vegetales (especialmente las hojas) por transpiración disminuyen la presión osmótica celular. A medida que el agua se mueve hacia arriba desde las raíces para reemplazar el agua utilizada y perdida por las células foliares, la presión osmótica disminuye en las células ciliadas de la raíz fina (con su alta área de superficie). Esto atrae agua hacia las células y raíces por ósmosis. Así, la presión osmótica es la principal fuerza que impulsa el agua hacia las plantas y, desafiando la gravedad, moviéndola desde las raíces hasta el resto de la planta
3. Osmosis en la vida animal
Los cambios en el ambiente osmótico pueden estresar o matar a un organismo. Por ejemplo, los organismos de agua dulce (protozoos o peces) colocados en el agua de mar morirán. De igual manera, peces de agua salada colocados en agua dulce. Pero los organismos pueden osmoregular (controlar la presión osmótica en sus células), al menos hasta cierto punto. Paramecio por ejemplo, expulsa el agua dulce para evitar que estalle a medida que toma agua. Esto se logra mediante una vacuola contráctil (mostrada a continuación).
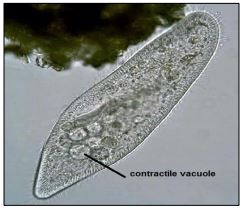
El agua ingresa constantemente a estos protistas de agua dulce porque la concentración de soluto en el citosol siempre es mayor que el agua dulce en la que viven. Para hacer frente a una captación constante de agua, sus vacuolas contráctiles recogen el exceso de agua y luego se contraen para expulsar el agua. Con un alto costo de energía, Paramecia constantemente bombea agua fuera de la celda para mantener el equilibrio hídrico (es decir, corregir la presión osmótica). Otra estrategia protista para hacer frente al cambio en las concentraciones ambientales de solutos (salinidad) es bombear sales (o sustitutos de solutos salinos adecuados dentro o fuera de la célula, según sea necesario (Para algunos detalles, ver Genes de Osmoregulación Protista Adquiridos por Eucariotas a partir de Bacterias por Transferencia Génica Horizontal).
¡Los organismos más grandes como los peces de agua dulce hacen frente a su entorno hipotónico orinando mucho! En el otro extremo del espectro, los peces de agua salada hacen frente a la alta concentración de solutos (sales) en su ambiente al excretar el exceso de sal. Los salmones pasan tiempo en agua de mar creciendo hasta la madurez y luego nadan río arriba en agua dulce para desovar. Se puede imaginar cómo el salmón y organismos similares tienen que osmoregular para adaptarse a sus entornos cambiantes, muy diferentes. En este caso, la osmorregulación comienza cuando los cambios hormonales responden a cambios en las circunstancias de vida y dictan una respuesta compensatoria.
4. Resumiendo
La ósmosis es el movimiento del agua a través de las membranas hacia donde los solutos están en alta concentración. Al mismo tiempo, los solutos que pueden difundirse a través de las membranas se mueven dentro o fuera de las células hacia donde se encuentran a menor concentración, ya sea pasivamente, o por difusión facilitada. Hemos desarrollado diferentes proteínas de transporte facilitado específicas para diferentes proteínas. Finalmente, la mayor parte del agua cruza las membranas al facilitar la difusión a través de proteínas de acuaporina que sirven como poros en las membranas celulares.
D. Transporte activo
La excitabilidad (adaptación) es otra de las propiedades definitorias de la vida. Esta propiedad de todas las células se basa en la reactividad química y eléctrica. Los neurotransmisores liberados en una sinapsis cruzan la hendidura sináptica de una neurona “de envío” a una célula que responde (otra neurona o una célula muscular). El neurotransmisor se une a receptores en la célula respondedora dando como resultado una despolarización de la membrana, un cambio rápido en la diferencia de potencial eléctrico a través de la membrana celular. Si bien las respuestas a los neurotransmisores ocurren en fracciones de segundo, todas las células responden, aunque no siempre tan rápidas como las neuronas o las células musculares. Los cambios en la polaridad de la membrana de cualquier célula dependen de concentraciones desiguales de iones dentro y fuera de las células. Las células en reposo suelen tener una mayor [K +] en el citosol y mayor [Cl -] y [Na +] fuera de la célula (abajo).
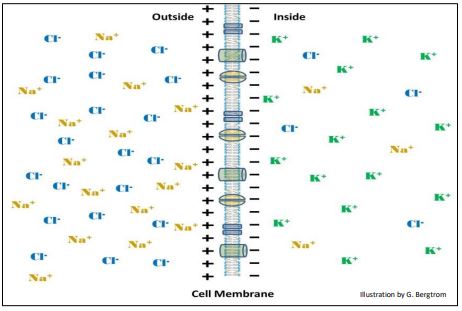
Estas diferencias iónicas a través de las membranas son las que permiten que células tales como las neuronas y los músculos respondan a señales químicas y otras señales (por ejemplo, eléctricas). Así, las células tienen un potencial de reposo, mostrado aquí con signos más y menos en lados opuestos de la membrana. El potencial de reposo medido (diferencia de carga o diferencia de potencial) de la mayoría de las células está típicamente entre -50mv y -70mv. La alteración del potencial de reposo (es decir, despolarización de la membrana), resulta de un flujo de iones a través de las membranas. Los potenciales de reposo sostenidos por gradientes iónicos permiten una respuesta fisiológica a señales químicas u otras.
Los potenciales de reposo cambian cuando las células se excitan, así como por la fuga de iones normal pero no funcional. Ya sea incidental o intencional, se debe restaurar y mantener el equilibrio iónico correcto. Esto se logra mediante el transporte activo de iones a través de la membrana. Este proceso energéticamente desfavorable requiere una entrada de energía libre, típicamente de la hidrólisis de ATP. La bomba Na + /K + es un complejo de proteína de transporte activo ligado a la actividad ATPasa. A continuación, consideramos el flujo iónico durante la excitación celular y cómo funcionan las bombas de iones.
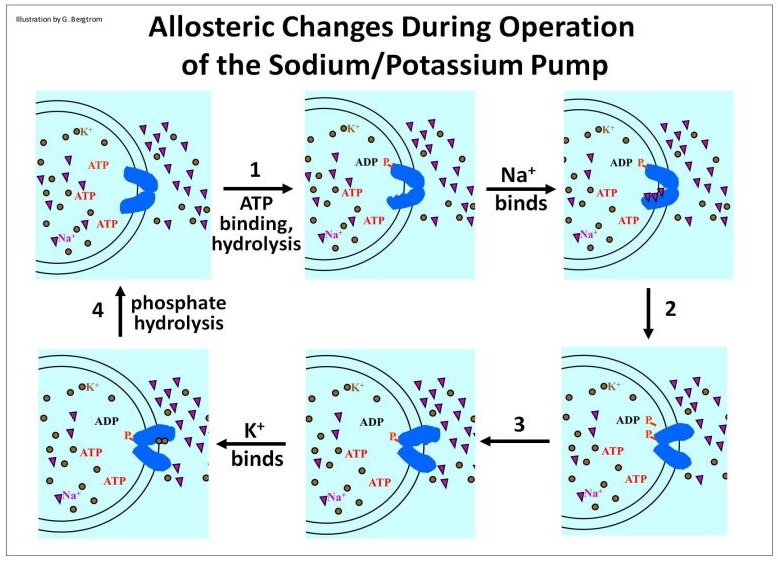
Comencemos por observar los cambios alostéricos que ocurren cuando la bomba de Na+ /K+ trabaja para restaurar y mantener gradientes de iones (ilustrado en la página siguiente). En funcionamiento, el dominio ATPasa de la bomba de Na+ /K+ hidroliza ATP, dejando un fosfato unido a la bomba e induciendo el primero de varios cambios alostéricos en las proteínas de la bomba (No. 1, anterior). En su nueva conformación, la bomba une tres iones Na+, provocando un segundo cambio conformacional que a su vez libera los iones Na+ en el fluido extracelular (No. 2). La liberación de iones Na+ fuera de la célula provoca un tercer cambio alostérico (No. 3), después de lo cual dos iones K+ del fluido extracelular son capaces de unirse a la proteína de la bomba. La unión de K+ provoca la hidrólisis del fosfato de la proteína de bombeo, devolviendo las proteínas de bombeo a su conformación original (No. 4) y liberando los dos iones K+ en el citosol. ¡La bomba Na+ /K+ está lista para la acción otra vez!
295 La fuga de potasio ayuda a mantener los potenciales de reposo celular
296 Transporte Activo por la Bomba de Sodio/Potasio
Por su descubrimiento de la bomba de sodio/potasio impulsada por ATPasa y sus estudios de cómo funciona para mantener el equilibrio de iones intracelulares, Jens C. Skou obtuvo una parte del Premio Nobel de Química 1997. Puedes leer más sobre Jens C. Skou en https://www.nobelprize.org/prizes/chemistry/1997/skou/auto-biography/