
17.5: Dirigiendo el tráfico de proteínas en las células
- Page ID
- 54458

Cada proteína polipeptídica traducida por ribosomas a partir de una secuencia de bases en un ARNm tiene una ubicación funcional específica, ya sea en el citoplasma, en membranas celulares, dentro de orgánulos o en fluidos extracelulares. En esta sección consideramos el movimiento y clasificación de proteínas en el sistema endomembrano así como el transporte de proteínas dentro y fuera de los orgánulos
A. Proteínas de envasado en el RER
Toda la síntesis de proteínas comienza de la misma manera, con la formación de un complejo de iniciación y los ciclos de elongación posteriores, formación de enlaces peptídicos y adición de aminoácidos carboxilterminales. Sin embargo, las proteínas secretoras y las destinadas a lisosomas, peroxisomas u otros microcuerpos, completan el alargamiento directamente en cisternas, o espacios encerrados por el retículo endoplásmico rugoso (RER). Es posible aislar y purificar proteínas secretadas por células cultivadas. Un buen sistema modelo para estudiar la síntesis secretora de proteínas resultan ser células de mieloma de ratón.
Se aislaron células de mieloma de ratón y se cultivaron en cultivo, donde sintetizan una cadena ligera de IgG, un polipéptido que forma parte de una molécula de inmunoglobulina de ratón. Las inmunoglobulinas se ensamblan a partir de polipéptidos de cadena ligera y pesada y se secretan a la circulación. Allí sirven como anticuerpos circulantes del sistema inmune vertebrado. Las células de mieloma de ratón son células cancerosas que han perdido la capacidad de producir los polipéptidos de la cadena pesada. En cambio, secretan principalmente la cadena ligera de IgG, lo que facilita su purificación del medio de cultivo celular. Un experimento temprano reveló que los polipéptidos secretados elaborados en un sistema de traducción in vitro son más grandes (más largos) que los mismos polipéptidos aislados de fluidos de secreción. Este experimento se resume en la página siguiente.
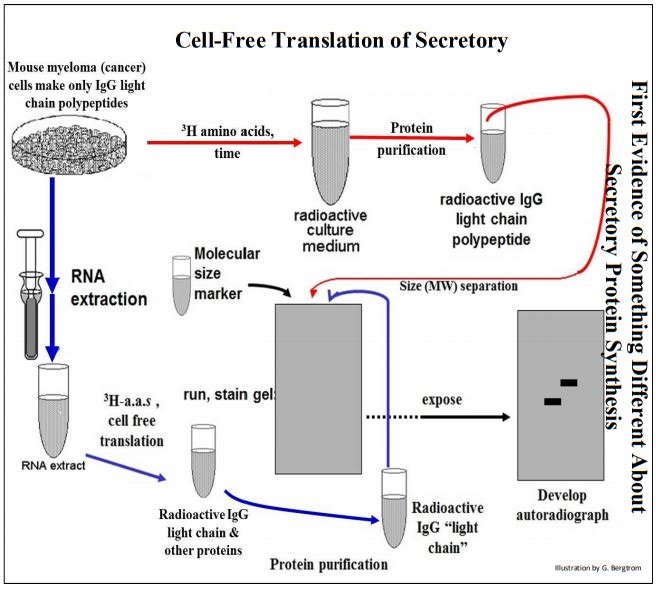
En una parte del experimento descrito anteriormente, las células de mieloma se cultivaron en presencia de aminoácidos radiactivos. Los polipéptidos de cadena ligera de IgG radiactivos resultantes se aislaron (siga las flechas rojas). El ARNm extraído por separado de otro lote de células de mieloma se agregó a un sistema de traducción libre de células que contenía aminoácidos radiactivos. Los polipéptidos radiactivos sintetizados in vivo e in vitro se separaron en geles electroforéticos y se autorradiografiaron (siga las flechas azules, arriba).
A partir de la autoradiografía, los polipéptidos maduros secretados hechos in vivo habían migrado más rápido en el gel que los traducidos in vitro. El producto de traducción libre de células fue de hecho, más grande que el polipéptido secretado maduro. Para explicar estos resultados, Gunther Blobel y sus colegas sugirieron la Hipótesis de la Señal, según la cual los genes de proteínas secretoras codifican aminoácidos adicionales como un péptido señal amino-terminal corto que dirige un polipéptido secretor en crecimiento hacia el RER. Para explicar la longitud más pequeña (es decir, más corta) del polipéptido maduro secretado, propusieron además que el péptido señal es solo una señal temporal de 'tráfico', eliminada por una enzima asociada a RER cuando el polipéptido cruzó la membrana del RER hacia el espacio cisterna.
304 Formulación de la hipótesis de la señal: experimentos tempranos
En la prueba de la Hipótesis de la Señal (que ganó Blobel el Premio Nobel de Fisiología o Medicina 1999), se incluyeron membranas RER aisladas con ARNm de células de mieloma de ratón en sistemas de síntesis de proteínas libres de células. Las autorradiografías electroforéticas mostraron esta vez que los polipéptidos elaborados in vitro en presencia de RER eran del mismo tamaño que los polipéptidos maduros secretados. Por lo tanto, el RER debe contener actividad de procesamiento, es decir, ¡una peptidasa señal que elimine el péptido señal! A continuación se ilustran los pasos de la hipótesis de la señal surgida de los experimentos de Blobel y sus colegas.
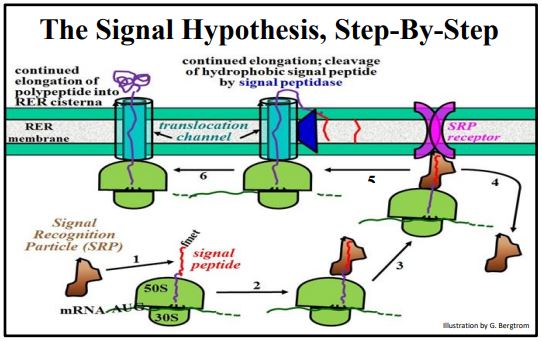
Recordemos que la síntesis de cualquier proteína comienza con el ensamblaje de un complejo de iniciación de la traducción, seguido por la elongación del polipéptido. Durante la elongación, el polipéptido en crecimiento se mueve y emerge de un canal, o surco en la subunidad grande. A medida que la secuencia señal N-terminal (es decir, el péptido señal) de un polipéptido secretor emerge de este surco, interactúa con la membrana RER. Comenzando en la parte inferior izquierda de la ilustración anterior, los pasos del proceso son:
- Una SRP (partícula de reconocimiento de señal) se une al péptido señal hidrófobo.
- La elongación se detiene hasta que el complejo SRP-ribosoma encuentra la membrana RER.
- El complejo ribosoma-SRP se une a un receptor de SRP en la membrana RER.
- La SRP se desprende de la cadena polipeptídica en crecimiento, para ser reciclada.
- El alargamiento de la traducción se reanuda a través de un canal de translocación; una peptidasa señal en la membrana del RER cataliza la hidrólisis co-traduccional del péptido señal, que permanece incrustado en la membrana del RER.
- El alargamiento continúa y el polipéptido en crecimiento comienza a plegarse en el RER.
305 Probando la Hipótesis de Señal
306 Detalles de la Hipótesis de la Señal
La etapa 2 anterior requiere que la SRP encuentre y se una al péptido señal antes de que el polipéptido naciente sea demasiado largo y comience a plegarse en una conformación 3D (terciaria). Resulta que el propio ribosoma puede mantener disponible el péptido señal desestabilizando interacciones electrostáticas que de otro modo conducirían al plegamiento y a una conformación indudablemente incorrecta. Para obtener más información sobre la participación de los ribosomas en el plegamiento de proteínas, consulte el enlace en Protein Folding-Desabilizing One Protein Strand at a Time.
El mecanismo secretor que se acaba de describir para los eucariotas tiene su contraparte en bacterias, que secretan proteínas que ayudan en la captación de nutrientes así como en la síntesis de la pared celular. Los péptidos señal parcialmente alargados guían los ribosomas unidos a ARNm hacia el lado citoplásmico de la membrana plasmática, donde los ribosomas se unen y luego pasan proteínas alargadas a través de la membrana plasmática hacia el espacio entre la membrana celular y la pared. A medida que la proteína sale de la célula, una peptidasa señal bacteriana (SPasa) escinde el péptido señal. Al parecer, el mecanismo para la secreción de proteínas evolucionó temprano y desde entonces se conservó. Como veremos, este mecanismo ha sido cooptado aún más por eucariotas para empaquetar proteínas en algunos orgánulos y en las propias membranas. Algunas especulaciones interesantes sobre la evolución de la ruta de empaquetamiento de proteínas se discuten en el siguiente enlace.
307 Destinos de Tráfico de Proteínas y Evolución de Caminos
Al principio, descubrimos que los antibióticos detienen el crecimiento bacteriano, ya sea interrumpiendo la pared celular o destruyendo las células directamente. Ahora sabemos que algunos antibióticos (por ejemplo, arilomicinas) alteran la función de la SPasa de la membrana plasmática, evitando que las proteínas requeridas en el espacio entre la pared celular y la membrana salgan de la célula. Una vez utilizadas contra Staphylococcus aurease, las arilomicinas ya no son efectivas porque muchas cepas se han vuelto resistentes a estos antibióticos (haga clic en Péptidasa de señal bacteriana y resistencia a antibióticos para leer sobre el mecanismo de resistencia a la arilomicina). Como ya sabrás, la S. aurease es ahora resistente a muchos antibióticos, y la enfermedad por infecciones intratables tiene su propio nombre, MRSA (Estaph Aureasa resistente a meticilina - cava por tu cuenta para ver más sobre la resistencia a la meticilina). Aunque se llama así por resistencia a la meticilina, el SARM ahora describe infecciones por S. aureasa casi intratables.
B. Síntesis de Proteínas (Integrales) que abarcan la membrana
Las secuencias señal N-terminales también guían a los ribosomas traduciendo proteínas integrales de membrana al RER. Sin embargo, antes de que dicha proteína pueda pasar completamente a través de la membrana, una secuencia de parada de transferencia (un dominio hidrófobo dentro de la cadena polipeptídica) atrapa la proteína en el interior de ácidos grasos de la membrana. Las múltiples secuencias de stoptransfer representan proteínas transmembrana que abarcan una membrana más de una vez (abajo).
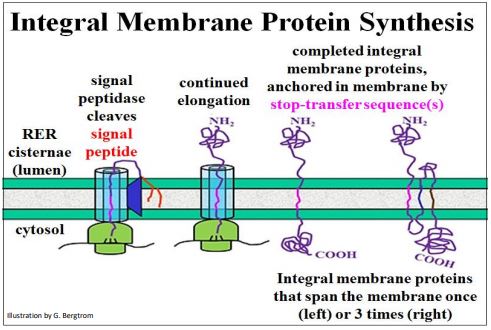
308 Proteínas Integrales de Membrana Secuencias de Transferencia
C. Traslado y clasificación de las proteínas envasadas a su destino final
Al igual que las proteínas empaquetadas en RER, las elaboradas en el citoplasma van a diferentes destinos antes de que se vuelvan funcionales. Veamos los mecanismos de clasificación de las proteínas secuestradas por el sistema endomembrano y las elaboradas en el citoplasma.
1. Tráfico en la Autopista Endomembrana
Ya hemos visto que, una vez empaquetadas en las cisternas del RER, las proteínas comienzan la modificación postraduccional (por ejemplo, por 'glicosilación nuclear'). Las vesículas transportadoras que brotan del RER transportan proteínas empaquetadas y de membrana a las vesículas cis del aparato de Golgi. Allí, la fusión de vesículas está mediada por el reconocimiento de proteínas integrales de membrana complementarias incrustadas en las dos membranas. Posteriormente, dichas proteínas empaquetadas se clasifican en diferentes orgánulos o a la membrana plasmática. La clasificación comienza a medida que las proteínas se mueven de la cara cis a la cara trans de las vesículas de Golgi, donde las proteínas de clasificación específicas se asocian con diferentes proteínas empaquetadas en las vesículas trans de Golgi. Las proteínas empaquetadas luego se clasifican en vesículas que brotan de las pilas trans de Golgi. Estas vesículas se desplazan a sus destinos finales, reconociendo y fusionándose con membranas adecuadas. Algunos eventos de tráfico de proteínas se animan en Eventos en Tráfico de Proteínas y se resumen en la ilustración de la página siguiente.
James E. Rothman, Randy W. Schekman y Thomas C. Südhof ganaron el Premio Nobel de Fisiología o Medicina 2013 por sus estudios de la regulación del tráfico de vesículas (haga clic en 2013 Premio Nobel de Fisiología o Medicina para más información). Vamos a seguir algunas proteínas dentro y sobre las membranas RER a través de la célula:
- Las vesículas de transición que llevan su mezcla de proteínas empaquetadas brotan del RER con la ayuda de las proteínas de la cubierta COPI y COPII, y se disocian de los ribosomas originalmente unidos a ellas. Sin embargo, las vesículas de transición permanecen asociadas con las proteínas COP.
- Estas vesículas se fusionan con las vesículas cis de Golgi, un proceso también mediado por proteínas COP. Las proteínas COCI se desprenden durante o después de la fusión, para ser recicladas de nuevo al RER.
- Las proteínas empaquetadas y las proteínas de membrana se procesan adicionalmente como paso a través de la pila de vesículas de Golgi, por ejemplo, sometidas a glicosilación terminal.
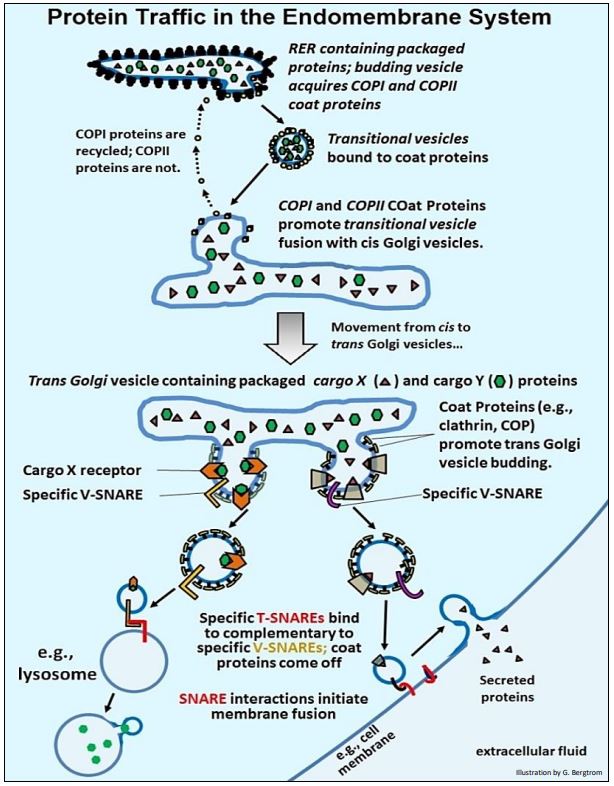
- En la cara trans de las vesículas de Golgi, las proteínas receptoras de carga en las membranas se unen a proteínas empaquetadas específicas (ahora llamadas proteínas de carga). Con la ayuda de clatrina y otras proteínas COP, las proteínas receptoras unidas a proteínas de carga brotan de la pila trans de Golgi. Sin embargo, esta vez, las proteínas de carga específicas se clasifican para separar vesículas con diferentes destinos celulares o extracelulares. Estas vesículas en ciernes también adquieren proteínas V-SNARE de membrana (para vesícula-SNARE).
- Cuando las proteínas V-SNARE en sus vesículas se unen a proteínas T-SNARE complementarias (para el SNARE objetivo) en las membranas receptoras, las membranas se fusionan.
- Algunas vesículas siguen esta vía, fusionándose con lisosomas o vesículas similares para almacenarlas con enzimas apropiadas y otro contenido proteico. Las proteínas de la cubierta salen de la vesícula fusionada y se reciclan, mientras que el contenido de la vesícula se transfiere a la siguiente vesícula.
- Las vesículas que contienen proteínas secretoras típicamente se fusionan para formar vesículas secretoras más grandes. Las vesículas secretoras pueden almacenarse hasta que se señale a las células que liberen su contenido de la célula. En ese punto, las vesículas de secreción se fusionan con la membrana plasmática, liberando su contenido al líquido extracelular. Una vez más, las proteínas de la cubierta y la clatrina se disocian de la vesícula secretora durante la fusión.
Otros jugadores se han quedado fuera de esta discusión, notablemente aquellos que hidrolizan trifosfatos de nucleótidos para proporcionar la energía para este tráfico de proteínas. Además, es posible que reconozcas a otros actores moleculares como la clatrina que desempeñan un papel en la endocitosis mediada por receptores. ¡Quizá eso no sea una sorpresa! Después de todo, la endocitosis es, al menos en parte, el tráfico molecular en la dirección opuesta a la formación y secreción de vesículas.
2. Tráfico de Proteína Nuclear
Casi todas las proteínas están codificadas en el núcleo y traducidas en el citosol. Estos incluyen la mayoría de los que se encuentran en el núcleo mismo, así como en las mitocondrias y cloroplastos (ver Hipótesis Endosimbiótica para una descripción de la expresión génica intraorganelar). Las proteínas sintetizadas en el citosol destinado a estos orgánulos contienen señales de tráfico de oligopéptidos que las dirigen a sus destinos apropiados. Anteriormente vimos que moléculas grandes (ARNm, ARNt) e incluso partículas enteras (es decir, subunidades ribosómicas) cruzan la envoltura nuclear a través de poros nucleares.
En cuanto a las proteínas dirigidas al núcleo, las señales de localización nuclear ricas en aminoácidos cargados positivamente (lisina, prolina) permiten la unión al dominio cargado negativamente de una proteína receptora de transporte nuclear en el citosol. Este proceso se ilustra a continuación.
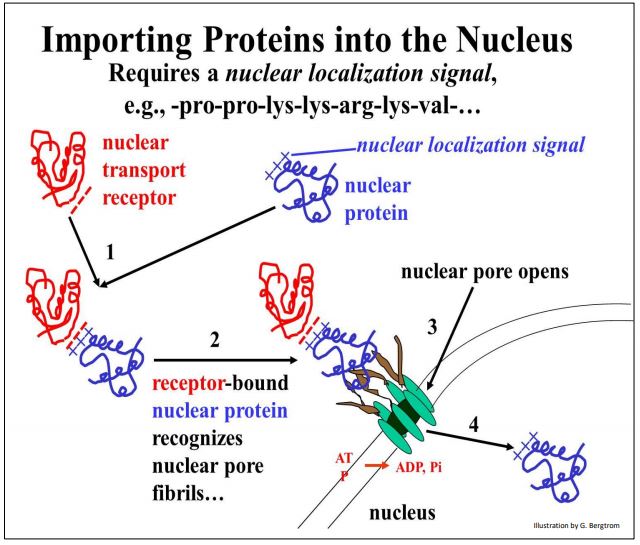
A medida que el complejo de las dos proteínas se acerca a un poro nuclear, interactúa con las fibrillas de poro nuclear, provocando que el poro se abra. Las dos proteínas unidas luego cruzan la doble membrana de la envoltura nuclear donde se acumulan contra un gradiente de concentración. Este transporte activo proviene de la hidrólisis de ATP a medida que las proteínas nucleares ingresan al núcleo.
3. Tráfico de Proteínas Mitocondriales
Recordemos que las mitocondrias contienen su propio genoma y maquinaria traslacional. Así, transcriben ARN y traducen proteínas propias. Sin embargo, los genes en el núcleo codifican muchas de las proteínas que se encuentran en las mitocondrias. La importación de estas proteínas a las mitocondrias se ilustra a continuación.
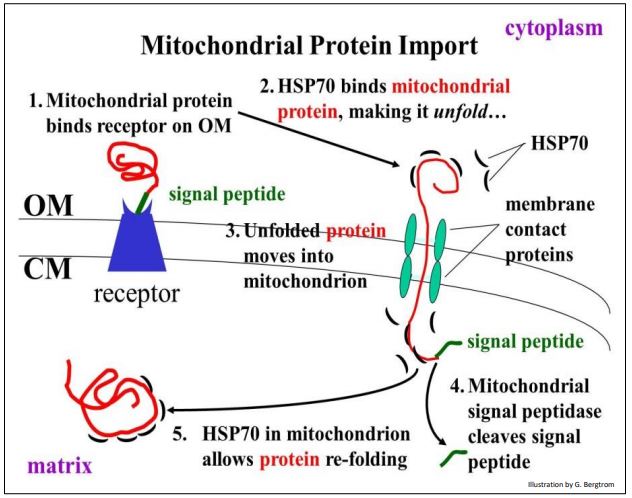
A diferencia del empaquetamiento co-traduccional de proteínas por el RER, la transferencia mitocondrial de proteínas es postraduccional. Esto significa que las proteínas mitocondriales formadas en el citoplasma ya se han plegado, asumiendo una estructura terciaria. Sin embargo, la proteína plegada expone un péptido señal N-terminal en su superficie que reconoce y se une a una proteína receptora en la membrana mitocondrial externa. La proteína receptora abarca tanto la membrana externa mitocondrial (OM) como la membrana cristalina (CM).
La proteína receptora entrega la proteína a proteínas de contacto con la membrana que también abarcan ambas membranas mitocondriales. Las proteínas de contacto con la membrana actúan como un canal, o poro, a través del cual la proteína mitocondrial cruzará hacia la matriz mitocondrial.
Pero hay un problema: ¡la proteína plegada no puede cruzar la membrana por sí misma! La entrada de una proteína mitocondrial completada en el citoplasma requiere una llamada proteína chaperona, en este caso la proteína HSP70 (choque térmico 70). HSP70 controla el despliegue de la proteína mitocondrial a medida que pasa a la matriz. Tras la eliminación del péptido señal por una peptidasa señal mitocondrial, otra molécula HSP70 residente en la mitocondria facilita el replegamiento de la proteína en una forma biológicamente activa. Recordemos que las HSP fueron descubiertas inicialmente en calor