
20.6: Charla de Moléculas- Selección de Comunicación Molecular y Complejidad
- Page ID
- 53889

En nuestra compleja sociedad humana, definimos la comunicación por su especificidad. Sin una cuidadosa elección de palabras, nuestro discurso sería en el mejor de los casos, una fuente de magníficos malentendidos, ¡o simplemente babel simple! ¿Qué significa esto para las químicas prebióticas?
En términos de evolución química prebiótica, la selección por definición habría favorecido la acumulación protectora de agregados moleculares de vida más larga. Con el tiempo, los mismos imperativos selectivos crearían redes de tales agregados, aumentando el rango y especificidad de las interacciones moleculares en un entorno desafiante. Si esto hubiera ocurrido en un espacio protocelular cerrado, debería haber resultado en una comunicación molecular primitiva y una complejidad creciente (¡otra propiedad de la vida!). De hecho, todas las propiedades de la vida deben haber acompañado el logro de una comunicación intermolecular cada vez más compleja. En pocas palabras, un cambio genético prebiótico (o para el caso un celular) que altere la velocidad de una reacción catalítica (si no destructiva) impulsará la selección de cambios en componentes de otras químicas metabólicas interconectadas. Si la comunicación molecular requirió la evolución de la especificidad catalítica, entonces la elaboración final de la complejidad y el orden como propiedad de la vida requiere además la selección de mecanismos de regulación y coordinación.
A. La comunicación intermolecular conduce a un establecimiento temprano de químicas interconectadas esenciales
Anteriormente, sugerimos que los precursores de catalizadores inorgánicos para enzimas biológicas probablemente eran minerales incrustados en arcilla u otros sustratos, proporcionando superficies que agregarían moléculas orgánicas de forma natural y catalizarían reacciones repetitivas. O bien los objetos iniciales de selección prebiótica incluyeron monómeros y polímeros estables externos, fuera o como parece más probable, dentro de proto-células. Posteriormente, la selección habría favorecido a los polímeros que potenciaron el crecimiento y reproducción de agregados exitosos. Estos polímeros fueron probablemente aquellos que catalizaron su propia síntesis, quizás colaborando con minerales catalíticos inorgánicos. El resultado sería la elaboración de una red de reacciones químicas interconectadas entre moléculas con alta afinidad entre sí, incrementando así la especificidad de esas reacciones. En el contexto de los orígenes y evolución de la vida, la co-catálisis describe las actividades de estas reacciones metabólicas interconectadas.
Como se señaló, las interacciones de alta afinidad son inherentemente protectoras. Durante la evolución química/metabólica prebiótica, los ensamblajes moleculares estables protegidos serían objetivos de selección. La continua coevolución de catalizadores, sustratos y conjuntos de reacciones co-catalíticas conduciría a una comunicación molecular cada vez más sofisticada. Una vez establecidos, los conjuntos de reacciones bioquímicas eficientes se verían restringidos contra cambios evolutivos significativos. ¡Cualquier cambio (mutación) que amenazara esta eficiencia significaría el fin de un linaje químico prebiótico (o para el caso, celular)! Esto explica por qué encontramos vías comunes para la generación de energía (por ejemplo, autótrofa y fermentativa), reproducción (replicación) y almacenamiento y recuperación de información (ADN, ARN, síntesis de proteínas) en todos los descendientes de LUCA.
La comunicación sofisticada y efectiva requiere coordinación. De hecho, la comunicación efectiva se define por la coordinación, la capacidad de tomar decisiones químicas. La selección de agregados moleculares que secuestraron reacciones metabólicas detrás de una membrana semipermeable asegura que solo ciertas moléculas se comuniquen entre sí. Es probable que este secuestro haya ocurrido repetidamente durante la evolución química, comenzando con la síntesis de moléculas poliméricas más grandes y posiblemente, una agregación de moléculas lipoidales primitivas. ¡Podemos pensar en la catálisis cada vez más efectiva en un ambiente cerrado como una conversación mediada por buenos oradores! ¡La coordinación es una propiedad que probablemente co-evolucionó con la vida misma!
B. Orígenes de la Coordinación
Veamos algunas estructuras posibles que se mueven alrededor en el conjunto de química prebiótica que podrían tener químicas compatibles de la vida autoensambladas o secuestradas. Junto con el compartimento de biopelícula de ventilación alcalina, los coacervados, las microesferas proteinoides y los liposomas han sido considerados como posibles progenitores de membranas biológicas. Cada uno se puede hacer en el laboratorio. Son demostrablemente semipermeables, ¡y en algunos casos incluso pueden replicarse! A continuación se muestran las micrografías y la producción de coacervados, microesferas proteinoides y liposomas

Oparin había propuesto que la acción de la luz solar en ausencia de oxígeno podría provocar que moléculas orgánicas ionizadas, cargadas opuestamente (e, g, aminoácidos, carbohidratos, etc.) formen gotitas a partir de moléculas orgánicas en su sopa primordial. Estos coacervados fueron producidos en 1932, visualizados por microscopía y demostrados como un compartimento semipermeable. Incluso se comportaron como si fueran capaces de crecer y reproducirse (también como Oparin sugirió originalmente que podrían hacerlo).
En la década de 1950, Sidney Fox produjo microesferas proteinoides a partir de péptidos cortos que se formaron espontáneamente a partir de soluciones acuosas de aminoácidos calentadas a sequedad (no muy diferente de lo que sucede en el escenario de charco de marea de formación de polímeros a partir de monómeros orgánicos). Estos se pueden ver por microscopía óptica y electrónica.
Si bien los liposomas se hacen fácilmente en un laboratorio, no está claro que existieran en una tierra prebiótica. Sin embargo, las membranas celulares deben haber adquirido su estructura bicapa fosfolipídica en la época de LUCA, ¡ya que todas las tenemos! Antes de LUCA, deben haber ocurrido rearrangenamientos químicos para permitir la incorporación de una bicapa de fosfolípidos en cualquier vida límite inicial con la que comenzó.
Ya hemos considerado la biopelícula propuesta para orígenes celulares en un respiradero alcalino. La formación de tales biopelículas habría separado protones ácidos oceánicos del interior de tales protocélulas, creando un gradiente de protones. Tal gradiente podría haber impulsado la evolución temprana de la quimiososis como medio para crear energía química, completa con la eventual selección de ATP sintasas y las enzimas de transporte de protones, nuevamente porque todas las células descendientes de LUCA poseen estas bioquímicas. Por supuesto, las microesferas proteinoides, coacervados, membranas basadas en biopelículas y liposomas no están vivas, y por lo tanto no son células. Pero uno u otro de ellos debió haber sido donde comenzó la coordinación potenciada de la comunicación molecular requerida para la vida.
354 Comunicación Molecular Protegida: Membranas Semipermeables
Un mensaje importante para llevar a casa aquí es que cualquiera que sea la estructura original de las primeras células, surgieron poco después de que los prerrequisitos químicos orgánicos de la vida comenzaran a adquirir funciones metabólicas familiares. Necesitamos ver el progreso químico y estructural hacia la celularidad como eventos evolutivos metabólicos concurrentes. En algún momento, la selección de bioquímicas secuestradas condujo a protocélulas, luego a las primeras células, cada una con todas las propiedades de la vida.
Finalmente, la selección de una comunicación altamente específica entre moléculas celulares permitió a las células hablar entre sí, participar en actividades grupales y finalmente unirse para formar organismos multicelulares. La multicelularidad es, por supuesto, una característica de muchos, si no de la mayoría, eucariotas. Pero vea una gran Charla TED sobre comunicación intercelular bacteriana a cargo de la Dra. Bonnie Bassler en Intercellular Communication in Bacteria.
C. Orígenes del almacenamiento y recuperación de información en un mundo de ARN
Aceptemos por ahora que la comunicación molecular comenzó simultáneamente con el empaquetado de conjuntos co-catalíticos interconectados en estructuras semipermeables. Luego se seleccionó la más “adecuada” de estas estructuras para una coordinación eficiente de mensajes químicos significativos y oportunos. En última instancia, la coordinación requiere procesamiento, almacenamiento y recuperación de la información, algo que reconocemos en el Dogma Central de Francis Crick del flujo de información del ADN al ARN a la proteína. Las células y los organismos hacen bastante bien la coordinación, pero ¿cómo son sus inicios? La respuesta puede estar en el mundo del ARN prebiótico que discutimos anteriormente. A continuación se muestra el Dogma Central, modificado para dar cuenta de la transcripción inversa y el comportamiento de los retrovirus.
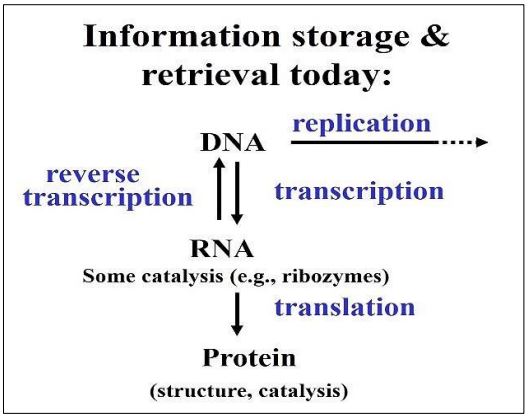
Realmente no sabemos cómo las células llegaron a depender del ADN para almacenar, transmitir y movilizar información genética, pero hemos presentado razones para creer que el primer ácido nucleico replicante fue el ARN, creando un mundo de ARN. Aquí está la evidencia que nos lleva a esta conclusión.
- Basándonos en el tallo y bucle y otras estructuras que se forman cuando las moléculas de ARN se someten a enlaces H internos, sabemos que los ARN pueden tomar formas variadas e intrincadas.
- Diversas conformaciones son consistentes con la evolución de la especificidad en la interacción de los ARN con ellos mismos y/o con otras moléculas en el ambiente prebiótico.
- Los ARN, ya sea solos como autocatalizadores (por ejemplo, ARNm de auto-corte y empalme) o en complejos catalíticos de ribonucleoproteínas (por ejemplo, ribosomas) existen en las células hoy en día.
- Algunos de estos ARN (específicamente ARNs), tienen una larga herencia filogenética, compartida por las células en los tres dominios de la vida.
La propensión de las moléculas de ARN monocatenario a plegarse basándose en la unión H interna puede conducir a esas diversas formas tridimensionales (estructura terciaria). Estas estructuras podrían haber interactuado con otras moléculas en un ambiente. Debido a que podrían replicarse de acuerdo con diferentes escenarios prebióticos, los mismos ARN también podrían transmitir información genética simple contenida en sus secuencias base. La combinación de propiedades informativas y catalíticas en una sola molécula se ilustra a continuación.
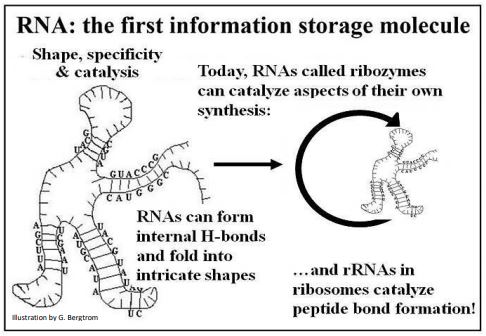
La capacidad de los ARN como catalizadores y almacenes de información genética habla de un candidato eficiente para el primer polímero dual o multipropósito, una propiedad que no se conoce y no se puede demostrar para el ADN. Lee más sobre los 'mundos de ARN' propuestos en los que la vida pudo haber comenzado en Cech TR (2012) [Los mundos del ARN en contexto. En Cold Spring Harbor Perspectivas en Biología (Cold Spring Harbor, NY: Cold Spring Harbor press) 4 (7) :a006742e].
355 Auto-Replicación: Información, Comunicación y Coordinación
¿Cómo podría haber sido la catálisis de ARN más allá de la autorreplicación en tiempos más simples? Considere la interacción entre dos ARNs hipotéticos y diferentes aminoácidos hipotéticos unidos a cada uno, que se muestra a continuación.
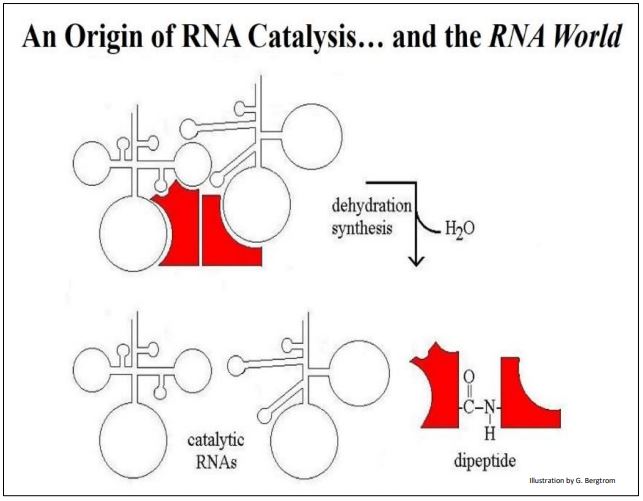
La unión de cada ARN a su aminoácido sería una interacción específica de alta afinidad basada en la complementariedad de carga y forma. De igual manera, los dos ARN que se ven en la ilustración deben tener una alta afinidad entre sí, también basados en complementariedades químicas y físicas. Incluso se puede imaginar algún fuerte enlace H entre bases en los dos ARN que podría desplazar la unión H intracatenaria (no se muestra aquí). El resultado es que los dos aminoácidos se unen de una manera que cataliza la formación de enlaces peptídicos. Esto requerirá una entrada de energía libre (recordemos que el enlace peptídico es una de las reacciones más intensivas en energía en las células). Por ahora, supongamos una fuente de energía química y centrémonos en las especificidades requeridas para la actividad catalítica del ARN.
Sabemos ahora que los ARNt son los intermediarios entre los ácidos nucleicos y la síntesis de polipéptidos. Por lo que es justo preguntarse cómo el tipo de actividad ilustrada anteriormente podría haber llevado a las interacciones ARNt-aminoácido que vemos hoy en día. No existe una química de unión obvia entre los aminoácidos actuales y los ARN, pero puede haber un legado menos obvio de las uniones propuestas. Esto tiene que ver con el hecho de que el código genético es universal, lo que significa que cualquier relación estructural entre ARN y aminoácidos debe haber sido seleccionada temprano (¡al inicio!) de la vida celular en la tierra. Aquí está el argumento.
- El código es de hecho universal (o casi así)
- Existe una correlación entre las propiedades químicas de los aminoácidos y sus codones, por ejemplo
- Los aminoácidos cargados (polares) son codificados por codones tripletes con más bases G (guanina).
- Los codones para aminoácidos no cargados contienen más a menudo una U media (uracilo) que cualquier otra base.
Estas correlaciones significarían que una unión temprana de aminoácidos a ARN específicamente plegados fue reemplazada en evolución por la unión covalente catalizada por enzimas de un aminoácido a un ARNt 'correcto', tal como vemos hoy.
¿Qué fuerzas podrían haber seleccionado la separación de las funciones combinadas de plantilla e información de la mayoría de las actividades catalíticas de los ARN? Quizás fue la selección de la mayor diversidad de estructura (es decir, forma) lo que pueden lograr los polipéptidos plegados, en comparación con los ARN plegados. Después de todo, los polipéptidos son cadenas de 20 aminoácidos diferentes en comparación con las cuatro bases que componen los ácidos nucleicos. Este potencial de diversidad molecular a su vez aceleraría el ritmo de la evolución química (y finalmente celular). Aquí se ilustra un escenario para la transición de eventos de ARN autorreplicantes anteriores a la traducción de proteínas a partir de ARNm.
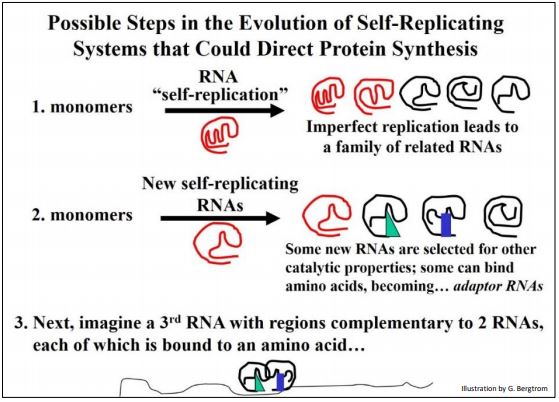
Los ARNs de adaptador en la ilustración se convertirán en ARNt. El nuevo ARN relativamente desplegado representa un ARNm presuntivo. Así, incluso antes de la entrada del ADN en nuestro mundo del ARN, es posible imaginar la selección de las características definitorias del código genético y mecanismo de traducción (síntesis de proteínas) que caracteriza a toda la vida en el planeta. A continuación, consideramos las “mejores especulaciones” sobre cómo el almacenamiento de información basado en ARN y las químicas catalíticas podrían haber hecho la transición evolutiva al almacenamiento de información basado en ADN y predominantemente catálisis enzimática basada en proteínas.
D. De Ribosomas a Enzimas; De ARN a ADN
El término co-catálisis podría muy bien describir reacciones bioquímicas en las que un catalizador acelera una reacción química cuyo producto retroalimenta de alguna manera en su propia síntesis. Esto lo vimos en acción cuando discutimos la regulación de enzimas alostéricas y el control de las vías bioquímicas. Los bucles de retroalimentación catalítica deben haber sido eventos significativos en la evolución de la comunicación intermolecular y la coordinación metabólica requerida para la vida.
Aquí consideraremos algunos escenarios para la transición de un mundo de ARN a algo más reconocible como el almacenamiento de información de ácidos nucleicos actual y el metabolismo catalítico basado en proteínas.
1. Rama de ribozimas: replicación, transcripción y traducción
Si los ARN catalizaron su propia replicación, puede haberse parecido a la replicación autocatalítica de AATE. Al mismo tiempo, algunos ARN también pueden haber atraído aminoácidos a sus superficies y catalizado la formación de enlaces peptídicos, como ya se describió. Por lo tanto, los ARN prebióticos bien formados pueden haber catalizado péptidos sintetizados, ¡algunos de los cuales eventualmente se harían cargo de la catálisis de la síntesis de ARN! El escenario se resume a continuación.
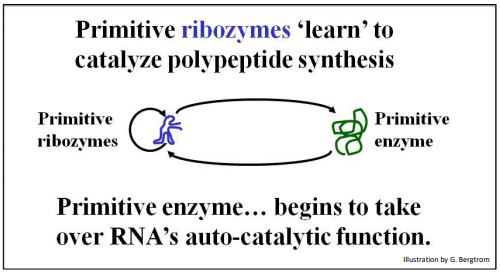
356 Almacenamiento y recuperación de información en un mundo de ARN
La selección que favorece la síntesis de oligopéptidos cortos y polipéptidos es consistente con una diversificación catalítica que condujo al dominio de catalizadores proteicos, es decir, enzimas. La enzima primitiva que se muestra aquí debió haber sido seleccionada porque al principio, ¡ayudó a la replicación autocatalítica del ARN mismo! Con el tiempo, la enzima evolucionaría junto con el ARN. Esta coevolución finalmente reemplazó la replicación de ARN autocatalítica con la síntesis de ARN catalizada por enzimas que reconocemos como transcripción hoy en día. En este escenario, ¡los preARNm y ribozimas autoempalmantes son remanentes de un mundo de ARN!
Pasemos ahora a algunas ideas sobre cómo un mundo de ARN podría hacer la transición al mundo ADN-ARN-proteína que tenemos hoy
2. Transferencia de Almacenamiento de Información de ARN a ADN
La transferencia de la función del ARN al ADN no es de ninguna manera un tema asentado entre los estudiantes de origen vital y evolución temprana. Una mejor suposición es que la elaboración de enzimas proteicas iniciada en el mundo del ARN conduciría a enzimas similares a transcriptasa inversa que copiaban información de ARN en moléculas de ADN. La información del ADN puede haber sido seleccionada porque el ADN es químicamente más estable que el ARN. A continuación se ilustra el escenario básico de transferencia.
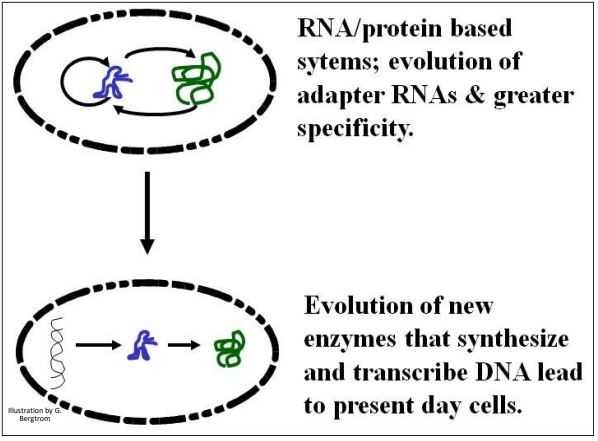
Todas las células vivas hoy almacenan información en el ADN (solo algunos virus tienen un genoma de ARN). Por lo tanto, la transición al uso del ADN como molécula de información debe haber precedido al origen de la vida. Al menos, debió haber ocurrido en las celdas de las que surgió el LUCA. ¡Los detalles de este cambio clave implican pasos evolutivos aún por resolver para satisfacción de todos!
E. La evolución de las vías bioquímicas
La historia de la evolución de las enzimas a partir de ribozimas y del ADN informativo del ARN, y las otras químicas metabólicas detrás de los límites semipermeables prebióticos está en curso en las células hoy en día. Sin duda, el metabolismo celular temprano implicó solo reacciones cruciales para la vida, catalizadas por un número limitado de enzimas. Pero, si la evolución tiende inexorablemente hacia una mayor complejidad de la comunicación y coordinación molecular, es decir, hacia una regulación cada vez más refinada del metabolismo, ¿cómo se hizo más grande el repertorio de enzimas y cómo se hicieron más elaboradas las vías bioquímicas? Respondimos a la primera pregunta en otra parte, cuando discutimos la duplicación de genes (por ejemplo, por cruce desigual). Los genes duplicados que codifican la misma enzima proporcionaron la materia prima para nuevas enzimas y nuevas funciones enzimáticas.
Ya sea en las células o en estructuras prebióticas, podemos plantear la hipótesis de cómo podría evolucionar una nueva reacción química. Por ejemplo, supongamos que una célula adquiere la molécula D requerida para una función esencial, de una fuente externa, ambiental. ¿Qué sucede si los niveles de D en el ambiente se vuelven limitantes? Claramente, las células morirían sin suficiente D. Es decir, a menos que las células que ya tengan un gen duplicado, redundante que haya mutado y que ahora codifique una enzima con la capacidad de hacer D en la célula. Tal célula podría haber existido con otras células sin la mutación, pero un entorno limitado en D seleccionaría la célula mutante para supervivencia y reproducción. Imagínese el escenario ilustrado a continuación.
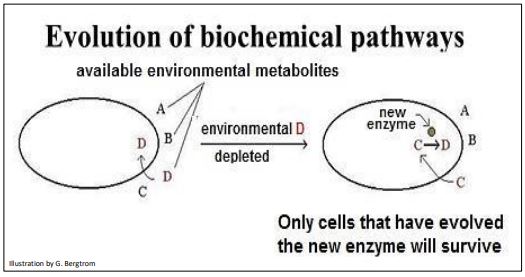
358 Orígenes y evolución de las vías bioquímicas
En un escenario similar, una mutación en un gen duplicado podría dar como resultado una nueva actividad enzimática que puede convertir alguna molécula (por ejemplo, C o D) en la célula en un nuevo producto molecular. Si la nueva enzima y el producto molecular no matan o debilitan la célula, la célula podría sobrevivir para ser seleccionada por alguna exigencia futura.