
1.2: Estudio de caso EvoDeVO
- Page ID
- 52779

Cuando miramos dentro de una especie vemos individuos altos y bajos (o grandes y pequeños) pero todos están escalados más o menos igual. Por ejemplo, generalmente no vemos personas altas con cuellos anormalmente largos o personas bajas con piernas anormalmente cortas. Como concepto general, podemos decir que este tipo de escalamiento debe ser el resultado de la intercomunicación entre diferentes células, tejidos y órganos. Por ejemplo, podríamos esperar que un hueso en crecimiento estire las fibras musculares e induzca a alargarse. También podríamos esperar que un hueso que crezca largo que también crezca amplio en proporción. Sin embargo, los mecanismos celulares y moleculares reales que realizan esta escala suelen ser algo misteriosos. Aquí vamos a comenzar a examinar un caso de descamación en un grupo de animales súper manipulables: los dípteros.
Bicoide en dípteros: Drosophila
Los dípteros son un tipo de mosca, y este grupo incluye las moscas con las que probablemente esté más familiarizado: las moscas domésticas y las moscas de la fruta de Drosophila. Como se mencionó en la Introducción, estas moscas experimentan un desarrollo extremadamente rápido. Esto es probablemente seleccionado por sus fuentes transitorias de alimentos. Además, sus embriones y larvas carecen de protección y son una buena fuente de nutrición para los depredadores insectívoros.
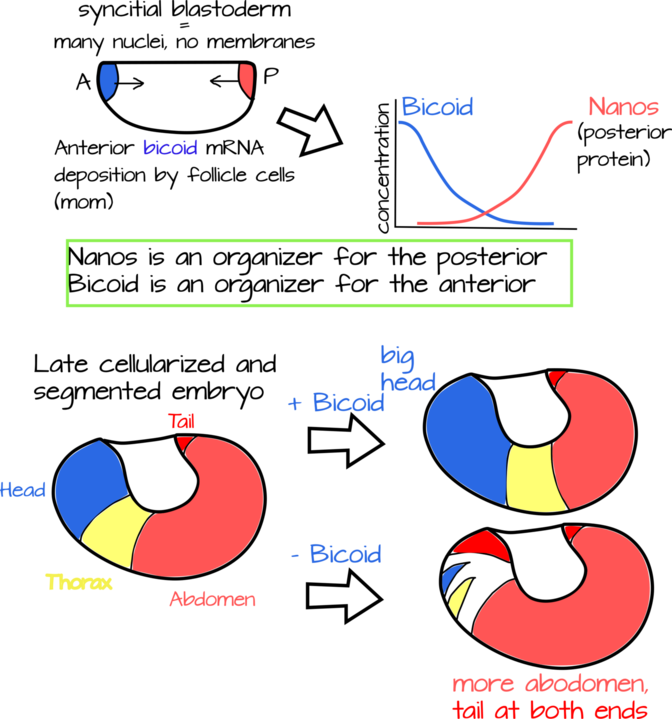
Una forma de acelerar el desarrollo es modelar a todo el animal al mismo tiempo en lugar de modelar primero un extremo, como hacemos nosotros y muchos otros animales. Drosophila y parientes (a menudo llamados los “dípteros superiores”) modelan sus extremos anterior y posterior al mismo tiempo. De hecho, el ovocito (óvulo) se modela incluso antes de que ocurra la fertilización, con el “organizador” del patrón anterior (Bicoide) en un extremo y el “organizador” del patrón posterior (Nanos) en el otro 1,2. Hay muchos más genes involucrados en este proceso, pero sobre todo nos centraremos aquí en Bicoid ya que ha sido objeto de intenso estudio.
Tanto los organizadores Nanos como los bicoides en Drosophila son proteínas que se encuentran en un gradiente de concentración. El bicoide tiene su mayor concentración en el extremo anterior, mientras que Nanos es mayor en el posterior. Hacia la mitad del embrión ambos están presentes solo en concentraciones bajas (Figura 2) 1,2,3,4 Si agotas Bicoide de un huevo de mosca, terminas con una mosca que le faltan estructuras anteriores (cabeza) y un abdomen expandido. Si agotas a Nanos, terminas con una mosca que le faltan estructuras posteriores (abdominales) y una cabeza expandida. Tenga en cuenta que la punta de la cabeza y la cola están modeladas por dos mecanismos de desarrollo separados, pero vamos a ignorar esto por ahora.
En este punto, voy a hacer nuestra primera incursión en la genética molecular ligera. Si te confundes un poco, por favor lee Brevemente, Genética para alguna aclaración.
Cuando Christiane Nusslein-Volhard hizo por primera vez mutantes Bicoides, se dio cuenta de un par de cosas interesantes. Primero, hay que hacer mutantes a las mamás. Esto tiene sentido porque es el ARNm materno el que se bombea al ovocito, así que por supuesto es el genotipo de la madre el que gobierna la morfología del embrión en este caso. Segundo, a pesar de que no tenía pruebas perfectas, notó que el gradiente de proteína Bicoid parece un gradiente de difusión. Un gradiente de difusión es el patrón que obtienes cuando algo se difunde libremente desde una fuente puntual (Figuras 2,3,4). Imagina que estás horneando galletas, un visitante llama a tu puerta. Al abrir la puerta, notan un olor débil pero fabuloso proveniente de tu casa. Hacen una pequeña charla contigo y caminan lentamente hacia la fuente del olor hacia la cocina. A medida que se acercan al horno el olor se intensifica. Abres el horno para sacar las galletas, y tu cocina se inunda con el olor de la bondad de las galletas. En este caso las galletas para hornear en tu horno son la fuente puntual y el olor ha formado un gradiente de difusión en tu casa, más débil en la puerta principal y más fuerte en la cocina. Este mecanismo para formar un gradiente se llama el “modelo SSD” para síntesis, soluto, difusión. No obstante, en el caso de Bicoid, este modelo tiene sus críticos, como descubriremos en El curioso caso del bicoide.

El problema del escalamiento en los dípteros
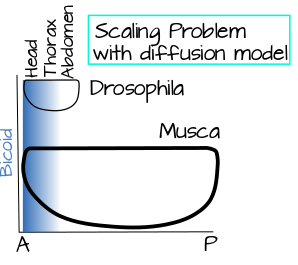
El modelo SSD se ve bien en papel, pero hay un dato confuso sobre los embriones dípteros que nos obliga a echarle un vistazo más de cerca. Los embriones dípteros varían enormemente en tamaño. Los embriones de Drosophila son pequeñísimos, de alrededor de un tercio a medio centímetro de largo. Los embriones de mosca doméstica, por otro lado, son la friolera de 1.5 centímetros de largo. ¿Qué significa esto para el modelo SSD? Imagina tu fuente puntual en nuestro ejemplo de galleta: un delicioso olor proveniente de tu horno. Sólo ahora estás cocinando en el comedor de tu Universidad. Alguien entra al comedor, ¿cuál es la probabilidad de que van a oler esas galletas? Bastante bajo, a menos que tengan un gran sentido del olfato. La puerta ahora está demasiado lejos para que el olor detectable alcance por difusión. De igual manera, en un gran embrión Musca (mosca doméstica), el gradiente de difusión solo alcanzaría 1/5 (¡o menos!) tan abajo del embrión como el mismo gradiente de difusión en un pequeño embrión de Drosophila (Figura 4). ¿Cómo resuelven las moscas este problema? Existen varias soluciones posibles que consideraremos en El curioso caso del bicoide.
Un archivo svg editable de la figura anterior se puede descargar en https://scholarlycommons.pacific.edu/open-images/5/
Referencias
- “La proteína bicoide determina la posición en el embrión de Drosophila de una manera dependiente de la concentración” 1988, Cell, Wolfgang Driever y Christiane Nüsslein-Volhard https://doi.org/10.1016/0092-8674(88)90183-3
- “Nanos es el determinante posterior localizado en Drosophila”, 1991, Cell, Charlotte Wang y Ruth Lehmann https://doi.org/10.1016/0092-8674(91)90110-K
- “De gradientes a rayas en la embriogénesis de Drosophila: llenando los vacíos”, 1996, Trends in Genetics, Rolando Rivera-Pomar y Herbert Jãckle https://doi.org/10.1016/0168-9525(96)10044-5
- “Formación y función del gradiente bicoide en el blastodermo presincitial de Drosophila”. 2016, eLife, Zehra Ali-Murthy y Thomas B Kornberg, 10.7554/elife.13222