
2: Fertilización y Rotación Cortical
- Page ID
- 52840

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Fertilización
Para las especies que se reproducen sexualmente, la finalización de la fertilización marca el inicio de la embriogénesis. Ya eres consciente de que las especies cuentan con diferentes mecanismos de aislamiento reproductivo que impiden la fertilización interespecífica. En el caso de los erizos de mar (enlaces a continuación) verás que hay varios pasos que pueden mediar en el reconocimiento específico de la especie. Estos incluyen quimio-atrayentes (sustancias químicas secretadas por el óvulo a las que responde el esperma) y moléculas pegajosas que contienen los espermatozoides (bindinas) que solo interactúan con los óvulos de la misma especie.
El punto de entrada de espermatozoides también es significativo en muchas especies ya que marca la primera ruptura en la simetría del huevo. Imagina una bola bastante simétrica. Ahora dibuja un solo punto en esa bola, este punto es ahora un hito. Puedes estar opuesto al punto, cerca del punto, o justo en el punto. El punto de entrada al esperma, al igual que el punto, actúa como un hito para romper la simetría. Lo hace de al menos dos formas relacionadas, donando un centrosoma al óvulo y desencadenando reordenamientos citoesqueléticos. El centrosoma espermático, un pequeño orgánulo que actúa como semilla para la formación de microtúbulos, puede actuar como organizador del aparato huso. Al poner un lado del huso cerca del punto de entrada al esperma, puede definir el eje de la primera escisión (Figura 1).
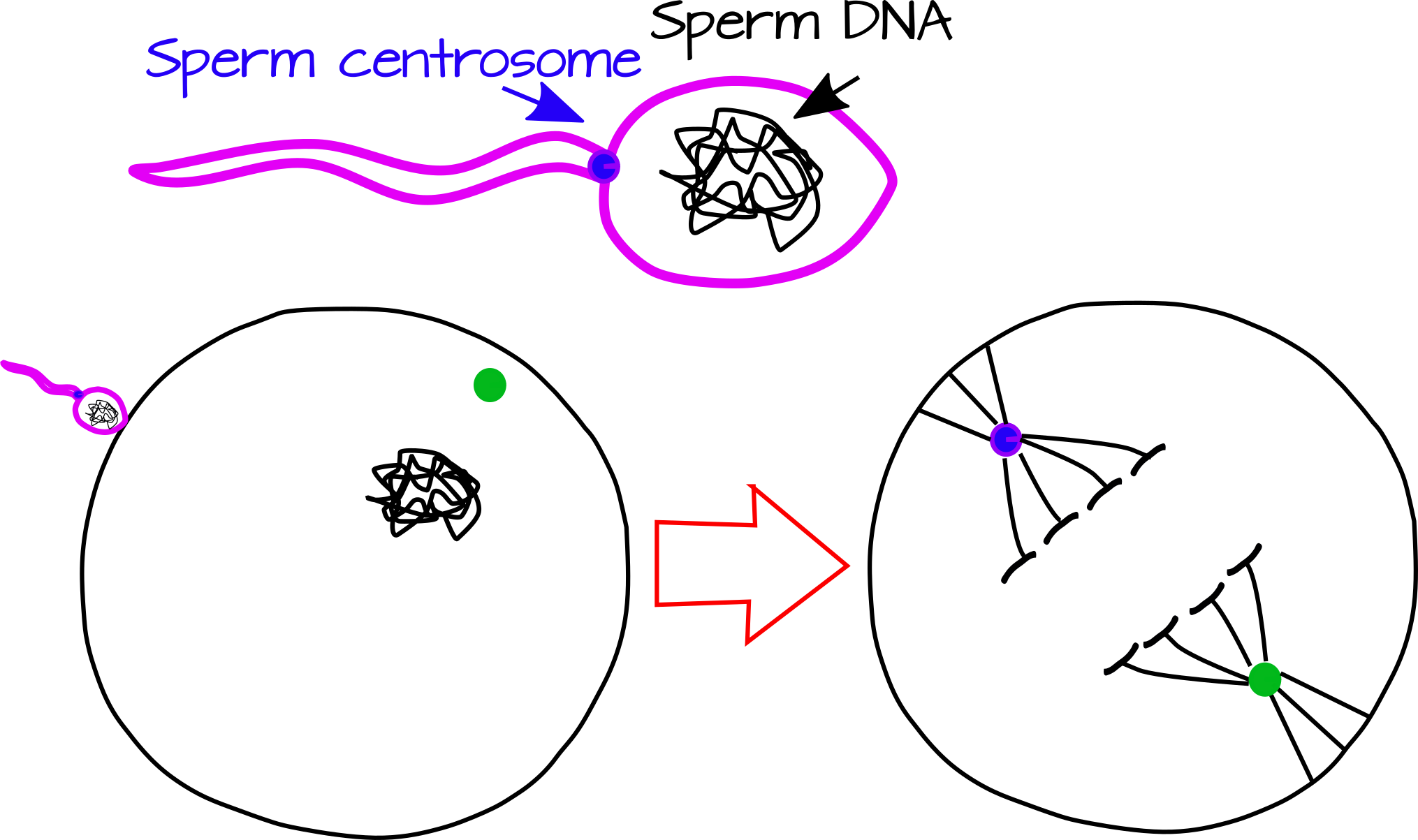
Adicionalmente, el punto de entrada al esperma puede ser un desencadenante del desplazamiento citoesquelético, incluyendo un proceso llamado rotación cortical, en el que la capa externa de la célula (la corteza) gira con respecto al citoplasma interno. Si bien la rotación cortical se limita a unas pocas especies (que conocemos), estudiarla ha llevado a avances en EvoDeVO y biología del desarrollo en general.
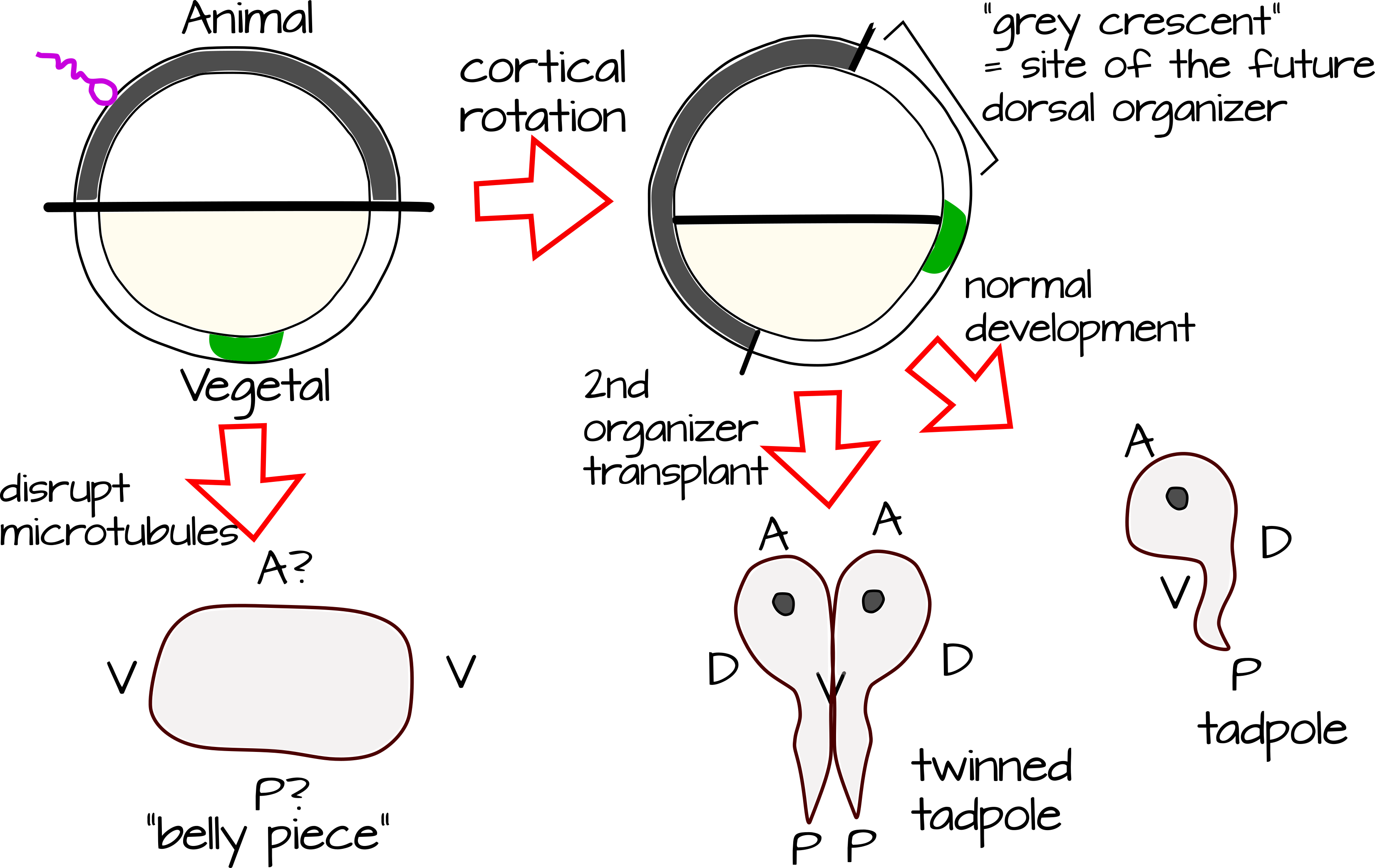
La importancia de la rotación cortical como evento de ruptura de simetría se subraya cuando vemos lo que sucede si lo evitamos. Los tratamientos breves de huevos fertilizados de rana (Xenopus laevis) con agentes que afectan la polimerización de los microtúbulos previenen la rotación cortical y dan como resultado un embrión “ventralizado”, es decir, tiene estructuras ventrales pero carece de dorsal estructuras incluyendo las que construirían una médula espinal. Llamamos a un embrión como este una “pieza de vientre”. ¿Qué tiene la rotación cortical que convierte a una rana en rana, más que en una masa de tejido del vientre?
Rotación cortical como ejemplo de EvoDeVO
A pesar de que la rotación cortical es un proceso que se encuentra en pocas especies animales, es un ejemplo de varios conceptos fundamentales en EvoDEVO
1) Establecimiento de organizadores. Los organizadores son partes del embrión que producen señales que modelan tejidos cercanos o distantes. En Drosophila, el ARNm bicoide marca el organizador principal, por ejemplo. El agotamiento de Bicoid conduce a moscas sin cabeza, demasiado Bicoid conduce a larvas de cabeza grande. Posteriormente en el semestre examinaremos al organizador ZPA en extremidades vertebradas.
Las ranas tienen un organizador Spemann-Mangold (que lleva el nombre de la estudiante Hilde Mangold y su asesor Hans Spemann que lo descubrió y caracterizó) 2. La extracción del tejido organizador conduce a una bola de tejido del vientre. La adición de un organizador a otra ubicación en el embrión conduce a renacuajos hermanados.
2) Inducción de la diferenciación celular. Muchas celdas tienen una ruta “predeterminada” que seguirán a lo largo del tiempo si no se les dan instrucciones adicionales. Una de las funciones principales de un organizador es hacer y enviar esas instrucciones adicionales. Los organizadores hacen esto a través de moléculas de señalización celular y regulación génica. El ejemplo bicoide en Drosophila es un ejemplo de regulación génica. El ejemplo de la rana demuestra tanto la regulación génica como la señalización celular, como pronto describiré.
3) El desarrollo ocurre en 4D. No sólo tenemos que pensar en objetos 3D complejos cuando estamos pensando en el desarrollo, sino que también debemos considerar el tiempo. Con el transcurso del tiempo, las celdas y partes de las celdas cambian sus posiciones. Esto cambia el “entorno local”, cambiando los tipos de interacciones que pueden ocurrir. Esto será más importante en la gastrulación, pero veremos el inicio de la misma aquí en rotación cortical. En el caso de la rotación cortical, la rotación cambia la ubicación de los ARNm y las proteínas. Estos eventualmente serán heredados por diferentes células y darán a estas células instrucciones de inicio para la diferenciación. Estas celdas luego cambiarán su posición y, a medida que entren en contacto con nuevos vecinos, se diferenciarán más y darán nuevas instrucciones.
La entrada de esperma desencadena cambios en el óvulo
Un ovocito de rana no fertilizado no es perfectamente simétrico. Tiene simetría radial, es decir se puede pegar un pin de arriba a abajo y será simétrico alrededor del pasador. Sin embargo, los lados superior e inferior difieren. El fondo (polo vegetal) se enriquece con gránulos densos de yema mientras que la parte superior (polo animal) se enriquece con citoplasma y pigmento menos densos. Similares simetría animal/vegetal se observa en muchas especies. Los huevos de rana también son asimétricos ya que están hechos de dos capas. Una corteza externa y una masa celular interna. Los polos animal y vegetal difieren en los ARNm y proteínas (ver Brevemente, Genética para un repaso en términos de biología molecular) 3 y también lo hacen las partes interna y externa del ovocito.
La entrada de espermatozoides induce un cambio en la forma en que los microtúbulos citoesqueléticos interactúan entre sí. Pasan de ser una red poco organizada a través de la célula a una matriz paralela con mayor densidad en la corteza y en el borde de la masa celular córtex/interior. Estos microtúbulos rotan la corteza con relación a la masa celular interna, llevando consigo los determinantes corticales localizados asimétricamente 2. Debido a que la pigmentación en el huevo se distribuyó de manera no uniforme, esto crea una “media luna gris” donde la corteza vegetal ahora se encuentra sobre la masa celular interna del animal (Figura 2)
La interacción entre los determinantes moleculares de la corteza vegetal y los determinantes moleculares de masa celular interna del animal hace que esta media luna gris sea única en comparación con el polo animal o vegetal del óvulo fertilizado. La media luna gris marca el futuro lado dorsal de la rana. Ahora se puede ver por qué la eliminación de la rotación cortical o la eliminación de los determinantes moleculares que pueblan la media luna gris conducen a la ventralización de la rana. La duplicación de la media luna gris, ya sea mediante trasplante o manipulación genética, conduce a un eje dorsal extra y al “hermanamiento” de la rana.
En el siguiente capítulo, Escote y Gastrulación, examinaremos el desarrollo y genética de este organizador y pensaremos en las consecuencias de organizar los tejidos en la evolución.
Referencias
- “Mitosis en el embrión humano: el papel vital del centrosoma espermático (centríolo)”, 1997, Histología e Histopatología, A. Henry Sathananthan PMID:
9225167 - “Inducción de primordios embrionarios por implantación de organizadores de una especie diferente”, 1924, Archiv für Mikroskopische Anatomie und Entwicklungsmechanik, Hans Spemann e Hilde Mangold., PMID: 11291841
- Revisado en: “Rotación cortical y localización del ARN mensajero en la formación del eje Xenopus”, 2012, WiRes Developmental Biology, Douglas W. Houston doi: 10.1002/wdev.29
Lectura adicional: Biología del desarrollo
www.ncbi.nlm.nih.gov/libros/nbk9983/
- Introducción a la fertilización
- 1. Esperma y Óvulo
- 2. ¿Cómo te va? Reconocimiento de esperma y óvulos
- 3. Un espermatozoide por óvulo: la reacción del gránulo cortical
- 4. OPCIONAL: La activación del metabolismo del huevo
- 5. OPCIONAL: Dos se convierten en uno, fusión de material genético
- 6. Rompiendo simetría: Reordenamiento del citoplasma del huevo
- 7. Resumen