3.3: El EvoDeVO del Blastopore: El Organizador Spemann-Mangold - Gastrulación en la Evolución
- Page ID
- 53073

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)El organizador Spemann-Mangold
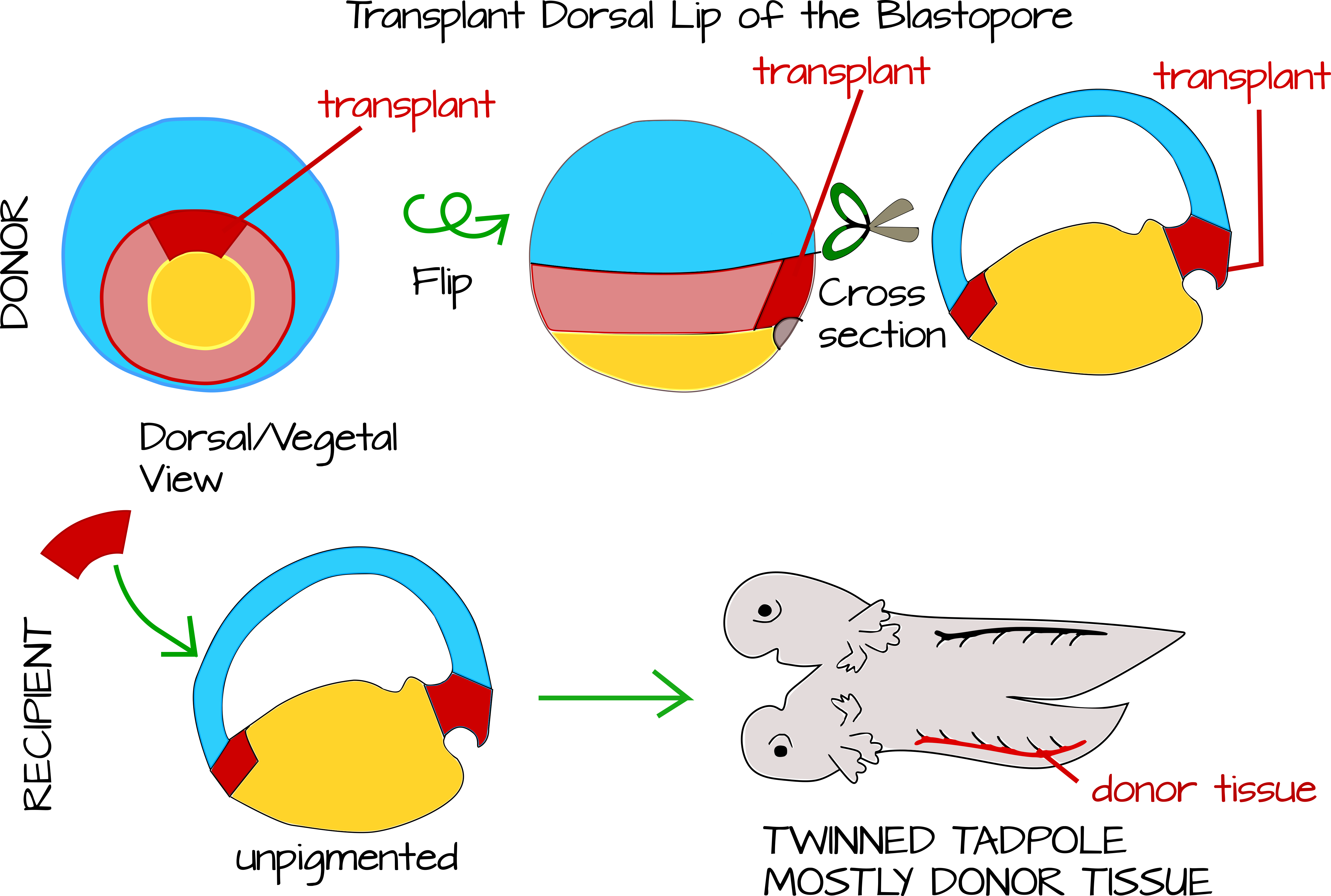
Hans Spemann y su estudiante de posgrado Hilde Mangold perfeccionaron una técnica para realizar trasplantes cruzados de especies del labio de blastoporo dorsal a nuevas ubicaciones en el cuerpo del huésped. Spemann se había preguntado durante años qué hacía exactamente el labio dorsal del blastoporo. Sabía que era importante porque si cortaba un embrión en dos, la mitad que obtuvo el labio del blastóporo dorsal se convirtió en un renacuajo, mientras que la otra mitad se convirtió en una “pieza de vientre”. También sabía que si dividía cuidadosamente el embrión por la mitad del labio dorsal del blastoporo, ambas mitades se convertirían en un renacuajo. Una posibilidad fue que una señal morfogénica emanara del labio dorsal que causara diferenciación celular en un gradiente de tiempo, de tal manera que las células más cercanas al labio dorsal se diferenciaran primero. Sin esta señal, las células no se diferenciarían. Spemann pensó que tenía apoyo para esta hipótesis cuando realizó experimentos de trasplante en tritones. Cuando trasplantó el labio dorsal de un tritón a otro, obtuvo tritones gemelos unidos, lo que sugiere que alguna señal del labio dorsal de hecho estaba modelando el eje del cuerpo principal 12,13.
Pronto Spemann revisó su hipótesis, basándose en gran medida en la observación cuidadosa de embriones gastrulantes y en el trabajo de trasplante de especies cruzadas de Mangold. Mangold tomó el labio de blastoporo de una especie de tritón no pigmentado y se trasplantó bajo el ectodermo ventral de un tritón pigmentado de gastrula temprana del mismo género. De esta manera pudo ver qué tejidos de renacuajo provenían del donante (el tritón no pigmentado) y cuáles del huésped (el tritón pigmentado). Encontraron que solo una pequeña porción del tejido de renacuajo hermanado estaba sin pigmentar, la notocorda (más algunas células adicionales aquí y allá, la técnica no era del todo perfecta). Esto significó dos cosas importantes: 1) El labio dorsal del blastoporo se desarrolla en la notocorda. 2) La notocorda modela el tejido alrededor de él para formar el eje primario del cuerpo 15.
Gastrulación en la evolución
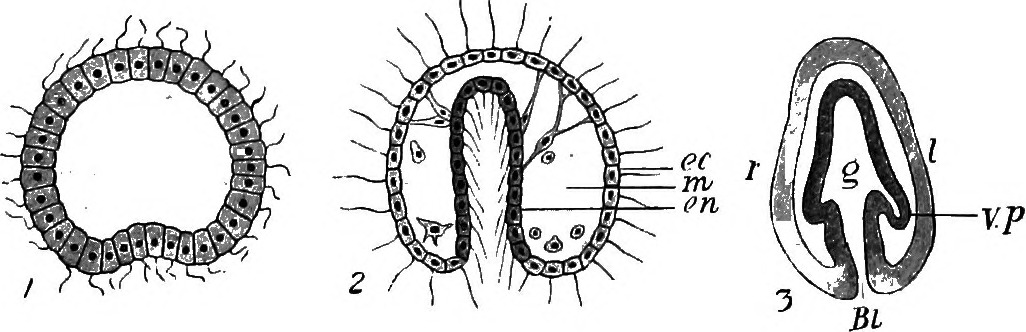
En general, la gastrulación implica la involución e invaginación de células de capa externa para internalizarse. Otras células de la capa externa se epibolizan para cubrir estas células internalizantes. De esta manera, un embrión pasa de un solo tipo celular a múltiples capas celulares distintas. El lugar donde ocurre esta invaginación o involución (donde las células se internalizan) se llama blastopore. El blastoporo puede ser un discreto “agujero” en el embrión que se alarga (llamado archenteron) como en el caso de los erizos de mar (Figura 7), o puede ser un labio ancho de células que se estrecha a medida que las células epibolizantes se expanden como en ranas y caracoles (Figuras 5 y 8).
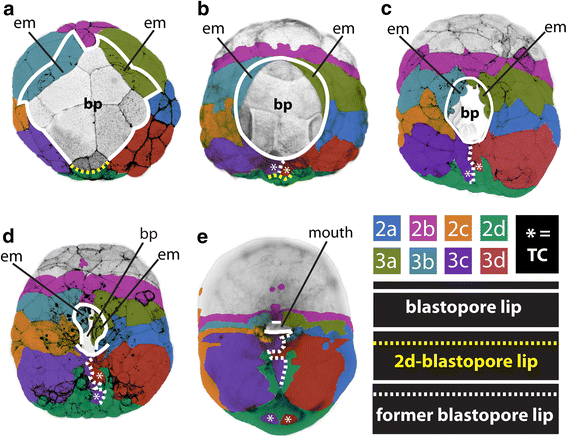
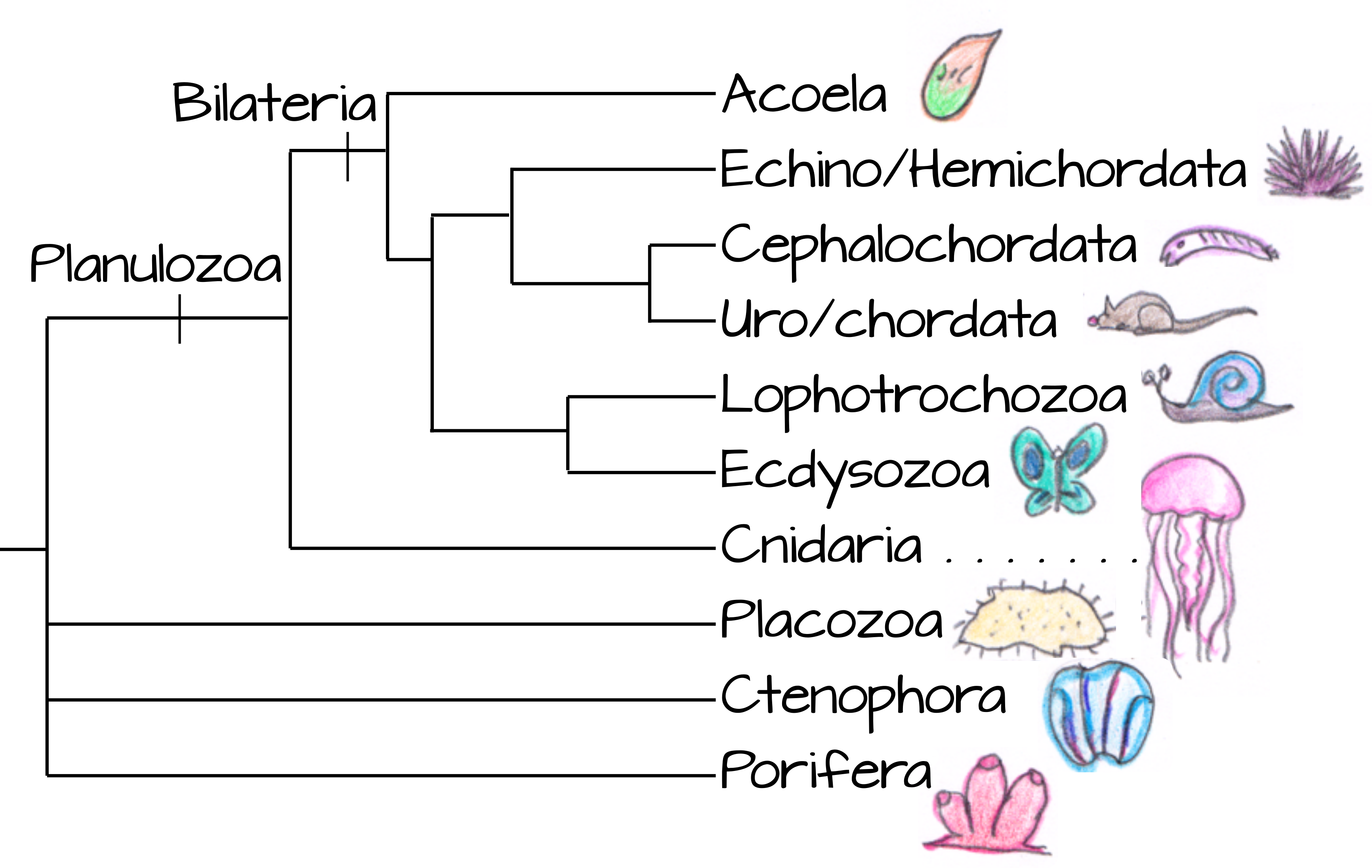
¿Cómo se pueden comparar e incluso homologizar dos estructuras tan diferentes? Consideramos que la gastrulación y la formación de blastoporas son homólogas a través de Planulozoa (el grupo que contiene la mayoría de los animales, Figura 9) por dos razones principales:
- Otros organismos multicelulares no sufren los reordenamientos masivos de tejidos que vemos en el desarrollo animal. De esta manera, la gastrulación representa una sinapomorfia animal (o al menos una Planulozoa).
- Hay puntos en común entre varios modos de gastrulación. El sitio inicial de gastrulación es casi siempre opuesto al sitio de extrusión del cuerpo polar, en el polo vegetal del animal 11. Y por supuesto el resultado final de la gastrulación -un tubo con ectodermo que recubre el exterior y un endodermo que reviste el interior con mesodermo o mesénquima en el medio- es claramente homólogo entre los planulozoos.