
9: Evolubilidad y Plasticidad
- Page ID
- 53133

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)La evolubilidad ha surgido como la conexión entre la evolución poblacional y la macroevolución. Aunque ambos procesos son fundamentalmente los mismos (ambos ocurren a través de mutación, deriva y selección), ocurren a escalas tan diferentes que normalmente usamos herramientas y mentalidades muy diferentes para estudiar cada uno de ellos. Por ejemplo, la evolución a nivel de población a menudo se modela con ecuaciones relacionadas con Hardy-Weinberg que examinan la aptitud y prevalencia de alelos particulares. Por otro lado, normalmente estudiamos el cambio macroevolutivo comparando distintas especies usando el registro fósil y la genética molecular. La pregunta es, entonces, ¿cómo vamos de aquí para allá? ¿Cómo estos procesos de micro-nivel eventualmente resultan en patrones de nivel macro? La evolubilidad es una forma útil de abordar esto. Una población es “evolutiva” si puede hacer frente a entornos cambiantes a través de la adaptación. Es decir, si las mutaciones genéticas en la población tienen buenas posibilidades de aumentar la condición física.
Si bien a nivel de superficie, esto parece simplista, podemos seguirlo a niveles más profundos para ver cómo la evolubilidad de una población afecta los procesos de desarrollo cuya evolución subyace a los cambios macroevolutivos. Gerhart y Kirschner abordan esto en su artículo Teoría de la variación facilitada. En este trabajo identifican las principales vías por las cuales los cambios genéticos del desarrollo pueden “facilitar” (o ayudar a aumentar) la variación fenotípica sin comprometer la aptitud (es decir, aumentan la evolubilidad).
Estas rutas, denominadas vinculación regulatoria débil, procesos exploratorios y compartimentación, se resumen a continuación:
Capacidad para participar en vínculos regulatorios débiles
La vinculación ocurre entre procesos que están conectados entre sí o con las mismas condiciones específicas. Por ejemplo, cuando tienes frío te tiemblas y te pones la piel de gallina. Estos dos procesos están conectados entre sí a través de una afección (frío) y también a través del sistema nervioso simpático. Asimismo, los procesos de desarrollo pueden vincularse a través de redes reguladoras génicas compartidas y/o a través de componentes reguladores compartidos (por ejemplo, genes que se expresan en respuesta al mismo factor de transcripción, o vías de transducción de señales que responden a la misma molécula de señalización).
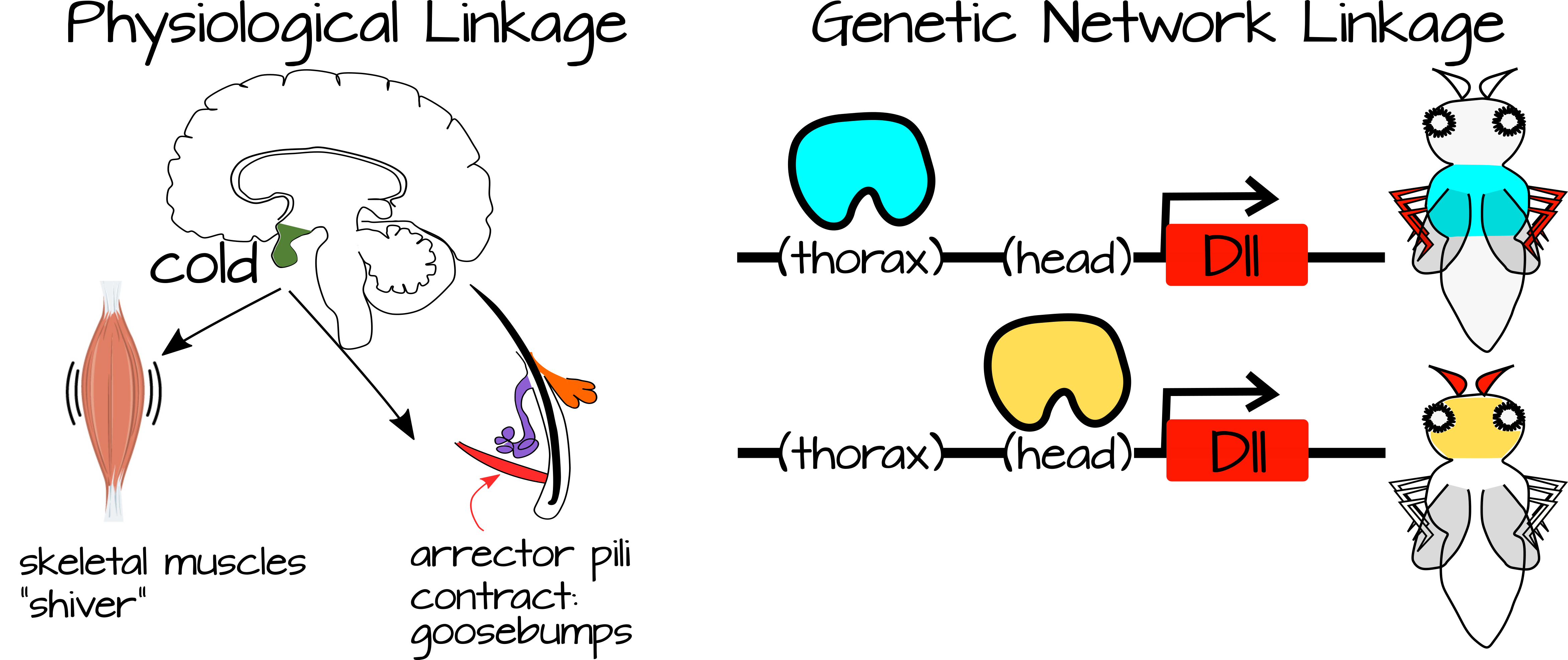
La débil vinculación regulatoria se refiere a procesos vinculados que están regulados por simples insumos que no proporcionan mucha información a los procesos. Hemos visto ejemplos de esto en Hábitos de vías de señalización altamente efectivas, donde un factor de transcripción libera represión sobre un conjunto de genes aguas abajo. Estos genes se autoactivan a través de activadores locales tan pronto como se libera la inhibición, lo que sugiere que potencialmente podría haber múltiples formas de desinhibirlos. Esto se considera un vínculo regulador débil porque la regulación por el factor de transcripción es “débil”. Cuando hay activadores locales, el valor predeterminado para el proceso es “on”. El factor de transcripción solo es necesario para liberar la inhibición. Una vez liberada esta inhibición, se puede producir una cascada de expresión génica, dando como resultado un proceso complejo con grandes consecuencias de desarrollo o fisiológicas. De esta manera se puede apagar o activar un proceso complejo en nuevos lugares con cambios regulatorios a pequeña escala. Por ejemplo, la expresión del factor de transcripción puede ser invertida hacia arriba o hacia abajo, u otras proteínas pueden actuar para modificar la capacidad del factor de transcripción para unirse a su ADN diana.
En general, la vinculación reguladora débil ocurre cuando una señal simple puede desencadenar múltiples procesos complejos dependiendo del contexto celular. Gerhart y Kirschner señalan que esto aumenta la plasticidad de un sistema ya que pequeños cambios en los factores reguladores (factores de transcripción por ejemplo) pueden cambiar la salida funcional de un sistema complejo. El complejo sistema en sí es en gran parte autorregulado, requiriendo solo un disparador de los factores reguladores. De esta manera, el desarrollo puede ocurrir más lentamente o más rápidamente en ciertas condiciones, las elecciones entre dos estados tisulares (por ejemplo, el tipo de gónada) pueden ser modificadas por factores ambientales, etc.
Procesos exploratorios
Los procesos exploratorios son procesos de búsqueda y búsqueda como seguir un gradiente de señal química a una fuente puntual. Vemos este tipo de procesos en desarrollo cuando examinamos cómo se forman los axones durante la neurogénesis y cómo surge la vasculatura para alimentar órganos y tejidos. Lo vemos en adultos en el sistema inmune adaptativo vertebrado y comportamiento de forrajeo en organismos guiados químicamente como hormigas. Decimos que este tipo de procesos muestra robustez porque se adapta rápidamente a los cambios ambientales locales. Por ejemplo, con un estricto patrón de la red XY de la vasculatura, el crecimiento a lo largo del tiempo de desarrollo tendría que estar bajo estricto control y no se toleraría la varianza de tamaño/forma en los órganos. Los vasos sanguíneos pequeños, sin embargo, no crecen debido a un sistema de coordenadas, sino que crecen en base a un sistema de “oferta y demanda” con excrecencias a regiones de bajo oxígeno. Las células secretan una señal proteica (VEGF) cuando son bajas en oxígeno, promoviendo el crecimiento de los vasos sanguíneos hacia ellas 2.
Los procesos exploratorios también son adaptables a lo largo del tiempo evolutivo ya que permiten la varianza de tamaño y forma. Esto es importante dentro de las poblaciones, donde los individuos pueden variar en tamaño o forma. Pero también es importante durante períodos más largos de tiempo evolutivo, ya que estos procesos flexibles de modelado formarán un sistema útil basado en su simple conjunto de reglas para el crecimiento. Por ejemplo, los discos de patas trasplantados en insectos se inervan al igual que las extremidades ectópicas en embriones de pollo 3 ,4. De esta manera, los procesos exploratorios facilitan la evolución al ayudar a construir un cuerpo viable cuando otros componentes morfológicos han evolucionado hacia conformaciones más ajustadas.
Compartimentación
En las plantas, la capacidad de separar las reacciones de luz del Ciclo Calvino de la fotosíntesis ya sea temporalmente (como en las plantas CAM) o espacialmente (como en las plantas C4) ha llevado a un éxito increíble y diversidad en estos tipos de plantas, ya que ya no son dependientes de mantener sus estomas abiertos durante el más caluroso y las partes más secas (pero también más soleadas) del día. En nuestras propias células, podemos ver la compartimentación de nuestras mitocondrias, con el espacio intermembrana aproximadamente 10 veces más ácido que el lumen. Esto permite que la fuerza motriz protónica se concentre y también protege el pH del resto de la célula. Nuestras células también separan físicamente la señalización célula-célula de la regulación transcripcional, lo que permite la modificación de las vías de señalización dependiendo de la historia celular (desarrollo) y el entorno.
Cuando un compartimento de un organismo (o célula) puede actuar de manera semiindependiente de otro, esperamos una mayor variación. Es decir, cada compartimento puede ejecutar su programa de desarrollo a través de la activación e inhibición de GRN específicos sin interrumpir el desarrollo de otros compartimentos. De esta manera, los cambios en la expresión génica que afectan solo a uno o unos pocos compartimentos podrían ser de efecto limitado. Por lo tanto, la evolución puede ser capaz de actuar sobre los compartimentos individualmente, siempre que estos sean cambios de expresión como el número de transcritos, isoformas de proteínas o modificación postraduccional. Las mutaciones codificadoras de proteínas, por otro lado, podrían actuar potencialmente en muchos compartimentos a la vez. La evolución puede actuar potencialmente en compartimentos individuales a través de cambios en la expresión génica, mientras que los procesos exploratorios pueden mantener la robustez de las vías de señalización y nutrientes en todo el cuerpo.
La evolubilidad en animales se basa en dos tipos de desarrollo competidores:
- Procesos resistentes al cambio evolutivo. Estos incluyen procesos exploratorios que utilizan señales externas para encontrar la morfología más eficiente o más efectiva, conduciendo a la retención del proceso de. Pero estos también pueden incluir procesos que se conservan durante largos periodos evolutivos de tiempo debido a limitaciones físicas o genéticas.
- Procesos cambiables. Estos incluyen procesos que cambian con el tiempo porque están compartimentados o cooptados en otros programas de desarrollo robustos. La compartimentación (un proceso resistente al cambio) permite la separación física de los procesos de desarrollo, y la débil vinculación regulatoria permite la cooptación de procesos a nuevos tiempos y lugares (por ejemplo, compartimentos).
De esta manera, el cambio evolutivo ocurre sobre un fondo de procesos robustos. Algunos de estos procesos robustos son maleables en el tiempo de desarrollo (como la vascularización), mientras que otros son limitaciones, limitando lo que es posible en un linaje evolutivo particular. Un ejemplo que ya hemos considerado es la evolución de las extremidades tetrápodas. Si bien existen limitaciones físicas y genéticas en esto que limitan el número de condensaciones óseas por sección de desarrollo, estas restricciones también crean compartimentos: el estilópodo, el zeugopodo y el autópodo, así como los 5 dígitos. La variación de tamaño y las adaptaciones a los compartimentos se sustentan en robustos procesos exploratorios que vinculan el desarrollo óseo con el apoyo de la musculatura, la vasculatura y la inervación.
Lectura adicional
Homología profunda (restricciones genéticas): https://www.sdbonline.org/sites/fly/lewhelddeep/deephomology.htm
Bauplans (limitaciones de desarrollo): https://evolution-outreach.springeropen.com/articles/10.1007/s12052-012-0424-z
La genética está modulada por el ambiente, esta modulación puede ser de corta duración (como broncearse) o de larga duración (como contraer diabetes insulinoresistente). Incluso puede atravesar generaciones (epigenotipo ahorrativo). ¿Cómo surge esta “plasticidad” en la genética y cómo cambia esto cómo funciona la evolución?
http://science.sciencemag.org/content/357/6350/eaan0221.long: Si haces tu genotipo 23andme, solo te dirá cuál es tu riesgo de ciertas enfermedades y trastornos. ¿Por qué no podemos estar seguros?
Referencias
- “La teoría de la variación facilitada” 2007, PNAS, John Gerhart y Marc Kirschner, https://doi.org/10.1073/pnas.0701035104
- Revisado en “Angiogénesis Inducida por Hipoxia: Bien y Mal”, 2011, Genes y Cáncer, Bryan L. Krock, Nicolas Skuli. y M. Celeste Simon doi: 10.1177/1947601911423654
- “Proyecciones axonales de patas ectópicas trasplantadas en un insecto”, 1985, Journal of Comparative Neurology, P. Sivasubramanian D. R. Nässel, doi.org/10.1002/cne.902390211
- “La inervación de extremidades adicionales inducidas por FGF en el embrión de pollo”, 2003, Journal of Anatomy, BW Turney, AM Rowan-Hull y JM Brown doi: 10.1046/j.1469-7580.2003.00131.x
- “Angiogénesis: un problema de modelado biológico dinámico adaptativo”, 2013, PLoS Biología Computacional, Timothy W. Secomb, Jonathan P. Alberding, Richard Hsu, Mark W. Dewhirst, Axel R. Pries https://doi.org/10.1371/journal.pcbi.1002983
- “Migración neuronal y laminación en la retina de vertebrados”, 2018, Fronteras en Neurociencia, Rana Amini, Mauricio Rocha-Martins, y Caren Norden. doi: 10.3389/fnins.2017.00742
La miniatura es una gráfica de variación de pie en una población (10,000 sitios polimórficos del cromosoma humano 1) de Graham Coop, publicada en Wikimedia Commons bajo licencia CC BY 3.0.