
10.2: Dactilia de aves
- Page ID
- 52873

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Las aves están en la clase Aves. Evolucionaron a partir de dinosaurios terópodos y el primer ave fósil descubierta fue Archaeopteryx. Algunas características que hacen de un animal un ave incluyen alas, picos desdentados y plumas. Las patas de pájaro tienen cuatro dígitos pero estos se pueden organizar de diferentes maneras. El arreglo de dígitos más común es tres en la parte delantera y una en la parte posterior como se ve en aves posadas y rapaces como el Halcón de la Sabana (Figura 1). La diferencia entre el alimento posado y el pie de rapaz no está en el diseño de dígitos sino en las garras. En el pie de rapaz, las garras son más curvas. También pueden tener dos en la parte delantera y dos en la parte posterior como se ve con el pájaro agarrador en la imagen. Otra variación en las patas de las aves son los dígitos palmeados que ocurren en aves nadadoras como patos.

Hay algunos arreglos de dígitos diferentes que se ven comúnmente en las patas de ave (Figura 2). Este arreglo más común se llama anisodactilia. Es donde los dígitos 2, 3 y 4 se enfrentan anteriormente y el hallux mira hacia atrás (Figura 2). Los pies de sindactilo tienen la misma disposición que el anisodáctilo, pero los dígitos 2, 3 y 4 están encerrados por la piel, esto se ve en las patas de martín pescador (Figura 1). También hay casos más extremos de cincha, especialmente en patos y otras aves acuáticas. Un pie de cigodáctilo es cuando dos dígitos, 2 y 3, están orientados anteriormente y los otros dos dígitos, 1 y 4 están orientados hacia atrás, un ejemplo de esto es en cucos y búhos (Figura 1). Un pie de heterodáctilo es similar a un pie de cigodáctilo pero los dígitos 3 y 4 miran anteriormente y los dígitos 1 y 2 miran hacia atrás, este pie es muy raro y solo se ve en trogones (Figura 1). Y finalmente un pie pamprodactyl es cuando los dígitos 1 y 4 pueden rotar para que los cuatro dígitos puedan enfrentar anteriormente. Los ratoneros y vencejos son famosos por sus pies pamprodactyl (Figura 2). Cuando sus dígitos se enfrentan de la misma manera, los vencejos (no en la foto) pueden usarlos como ganchos.
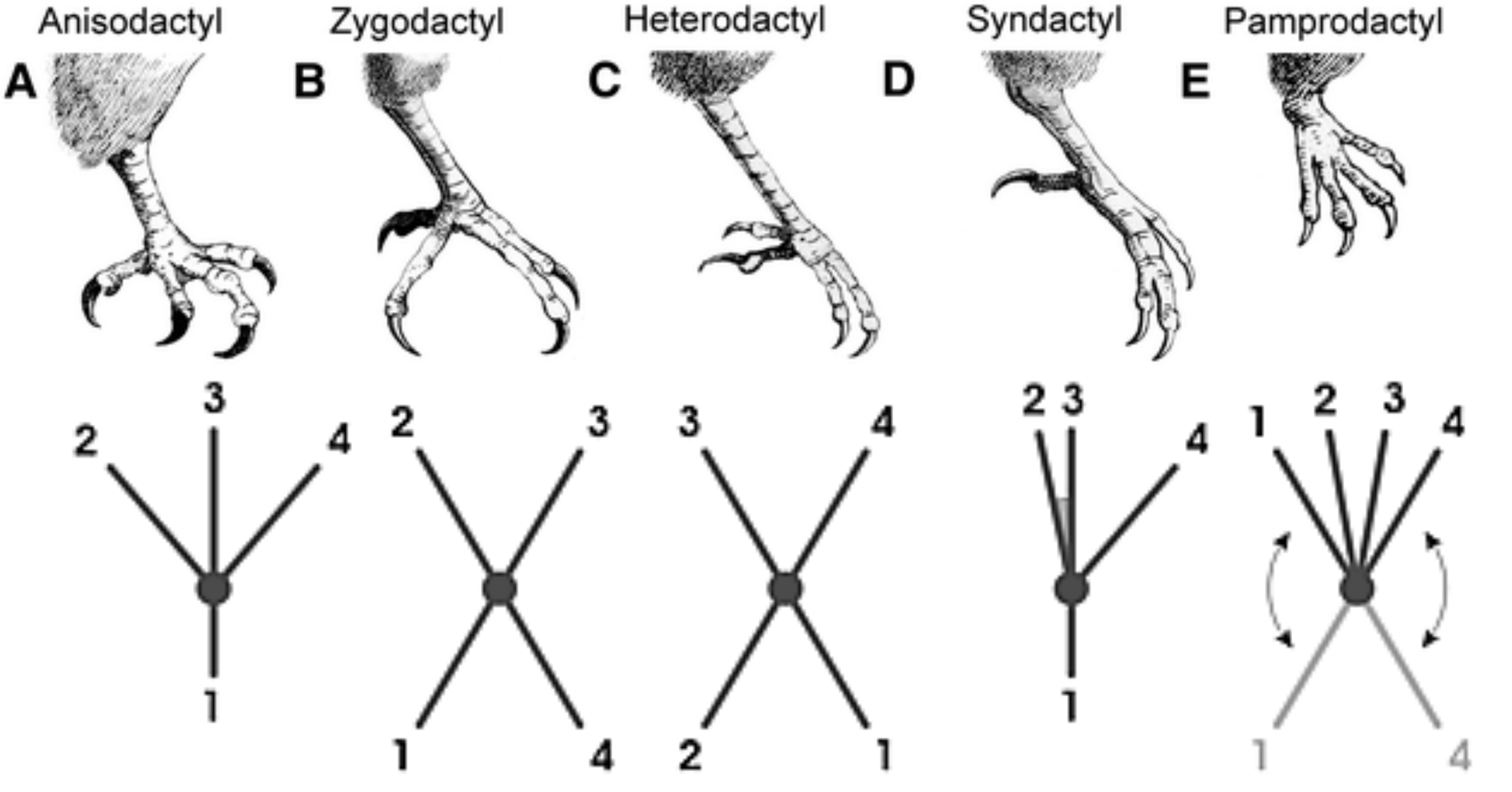 |
Figura 2: Arreglos de dígitos en patas de ave. El panel superior muestra dibujos lineales de patas de ave de Bothelo et al, 2015. Usado con permiso. El panel inferior muestra una “huella” de cada uno de los diferentes tipos de pies con dígitos numerados. Panel inferior de Wikimedia commons (Darekk2) y publicado originalmente bajo una licencia CC-BY-SA 3.0. |
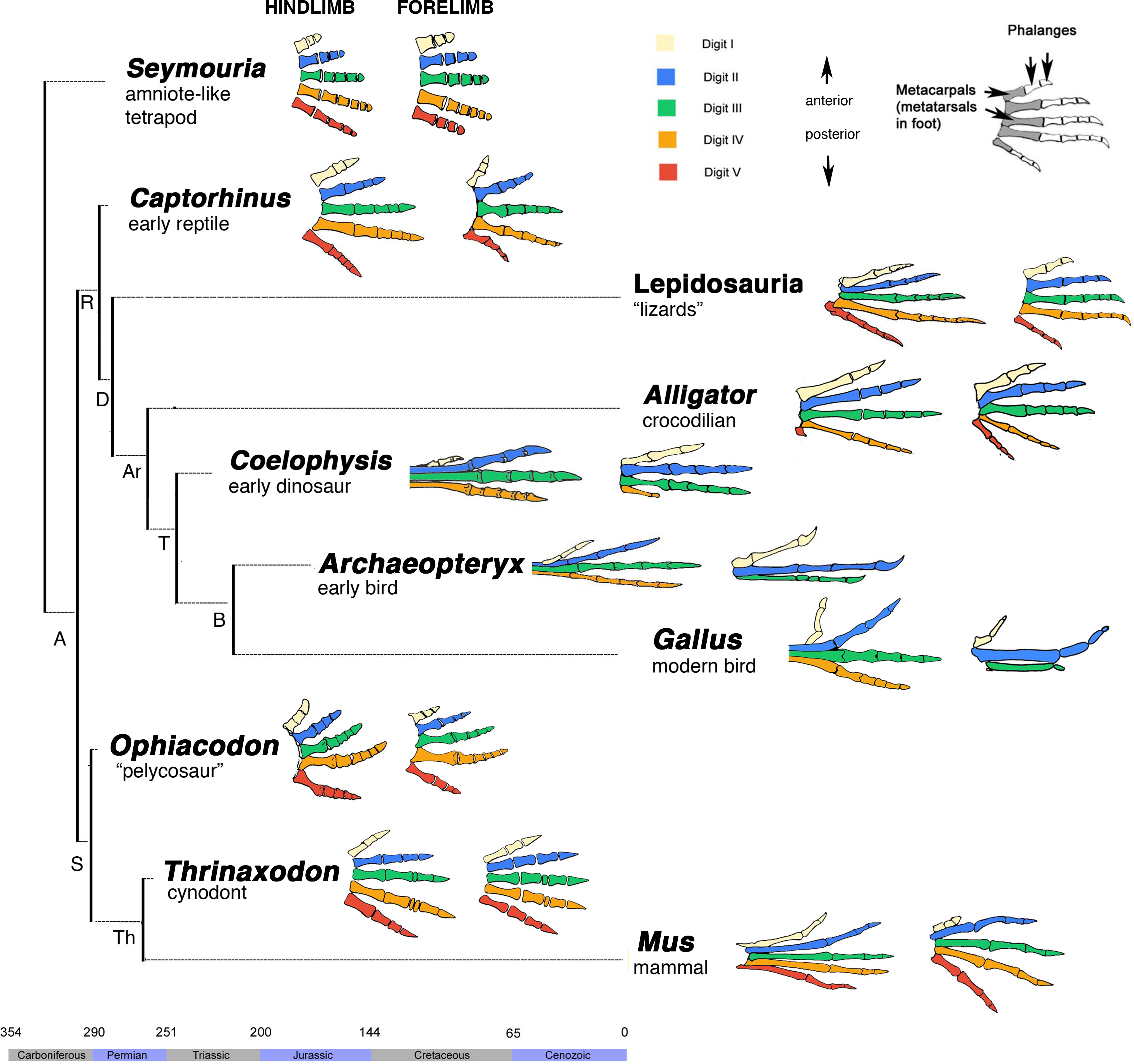
Las aves surgieron de dinosaurios terápodos con pies de 4 dedos (Figura 3). El pie de anisodáctilo se formó cuando el hallux se giró posteriormente. También solía estar ligeramente elevado y más pequeño que los otros dígitos. Ahora ya no está elevado y más largo que el segundo dígito. Esto dio como resultado los dígitos 2, 3 y 4 orientados hacia arriba y el dígito 1 posteriormente como se muestra en las imágenes anteriores. Se trata de una versión modificada del pie ancestral con pocos cambios. El pie de cigodáctilo se formó cuando ambos dígitos 1 y 4 giraron posteriormente (Figura 4). Este es un cambio más drástico que el pie de anisodáctilo.
 |
 |
Figura 3: Evolución de la morfología del dígito. Panel izquierdo: Los dígitos homólogos están codificados por colores. Las extremidades anteriores están a la derecha y las extremidades posteriores a la izquierda. La mayoría de los vertebrados tienen cinco dígitos en cada autopod, pero el linaje reptil muestra pérdida asimétrica de dígitos. Las aves, como Archaeopteryx y el polluelo (Gallus gallus) tienen cuatro dígitos en sus extremidades posteriores y tres en sus alas. Dentro del linaje de aves han evolucionado diferentes arreglos de dígitos. Imagen de Vargas et al, 2008 publicada bajo licencia CC BY. Panel derecho: la evolución de los arreglos de dígitos dentro del linaje de aves. Se desconoce el orden en que cada tipo de dactilia evolucionó y al menos una ha evolucionado varias veces. Figura de Jessica Niccum, Alexis Amador y Ajna Rivera
Botelho et al (2014) examinaron el desarrollo del pie de cigodáctilo visto en aves como periquitos. En este estudio, encontraron que en las etapas iniciales, los periquitos tuvieron un desarrollo similar al de un pollo (que tiene pies de anisodactilo). A diferencia de los pollos, sin embargo, después de aproximadamente 35 horas, el cuarto dígito giró mediolateralmente Después de 36 horas, el cuarto dígito se flexionó parecido a un pie de semi-cigodáctilo, como lo han hecho los búhos. Después de 37 horas, el cuarto dígito giró más para parecerse a la orientación del pie de cigodáctilo (Figura 4). Esto sugiere que la rotación de dígitos cerca del final del desarrollo del pie es responsable de la evolución de la cigodactilia, y los pies de zigodáctilo no crecen dígitos en una nueva ubicación. Debido a esta diferencia en etapas tardías, los investigadores examinaron la posibilidad de que la formación y activación muscular fuera responsable de la rotación de los dígitos.
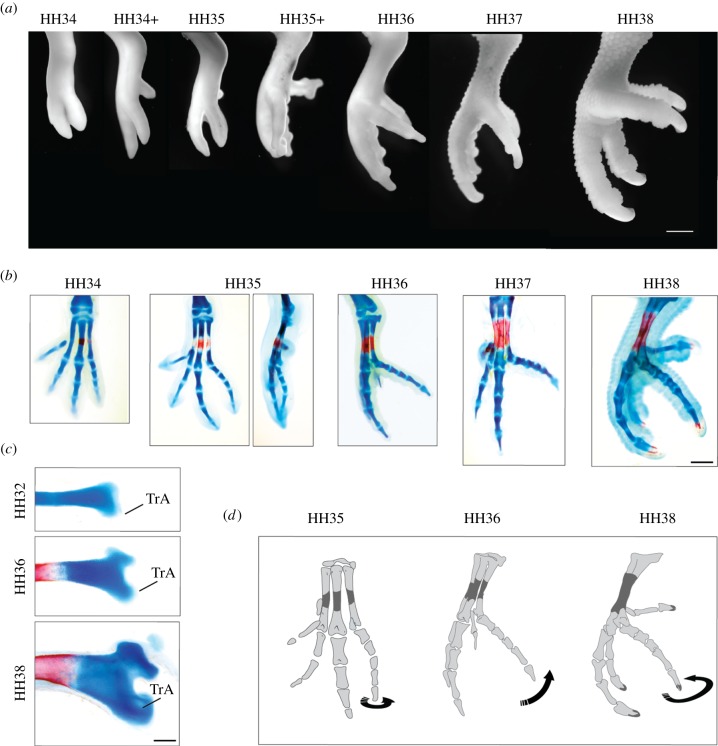 |
Figura 4: Desarrollo de un pie de cigodáctilo. A. Anatomía externa de un pie periquito en desarrollo en etapas sucesivas (HHXX). B. Estructuras esqueléticas en el desarrollo del pie periquito, el cartílago se tiñe de azul y los huesos rojos. C. trA es la trochlea accessoria, la punta distal del metatarsiano 4 La forma del anzuelo vista en HH38 es característica de las aves zigodáctiles. D. Diagrama del cambio en la orientación del dígito 4 durante el desarrollo. Figura de Botehlo et al 2014. Usado con permiso. |
Gran parte del desarrollo de diversas conformaciones de dígitos está ligado al desarrollo de la formación y activación muscular en las primeras etapas. En los pies de anisodáctilo, tanto EBDIV como ABDIV (músculos brevis extensores y aductores) están bien desarrollados. En cigodactilia como los periquitos, sin embargo, el EBDIV está casi completamente ausente. Los investigadores encontraron que el EBDIV se reduce cuando se separa de los otros músculos en los pies de cigodáctilo. El ABDIV, sin embargo, está bien desarrollado. El ABDIV provoca la flexión del 4to dígito. Esta flexión combinada con la restricción de EBDIV da como resultado la orientación del cigodáctilo (Figuras 4 y 5).
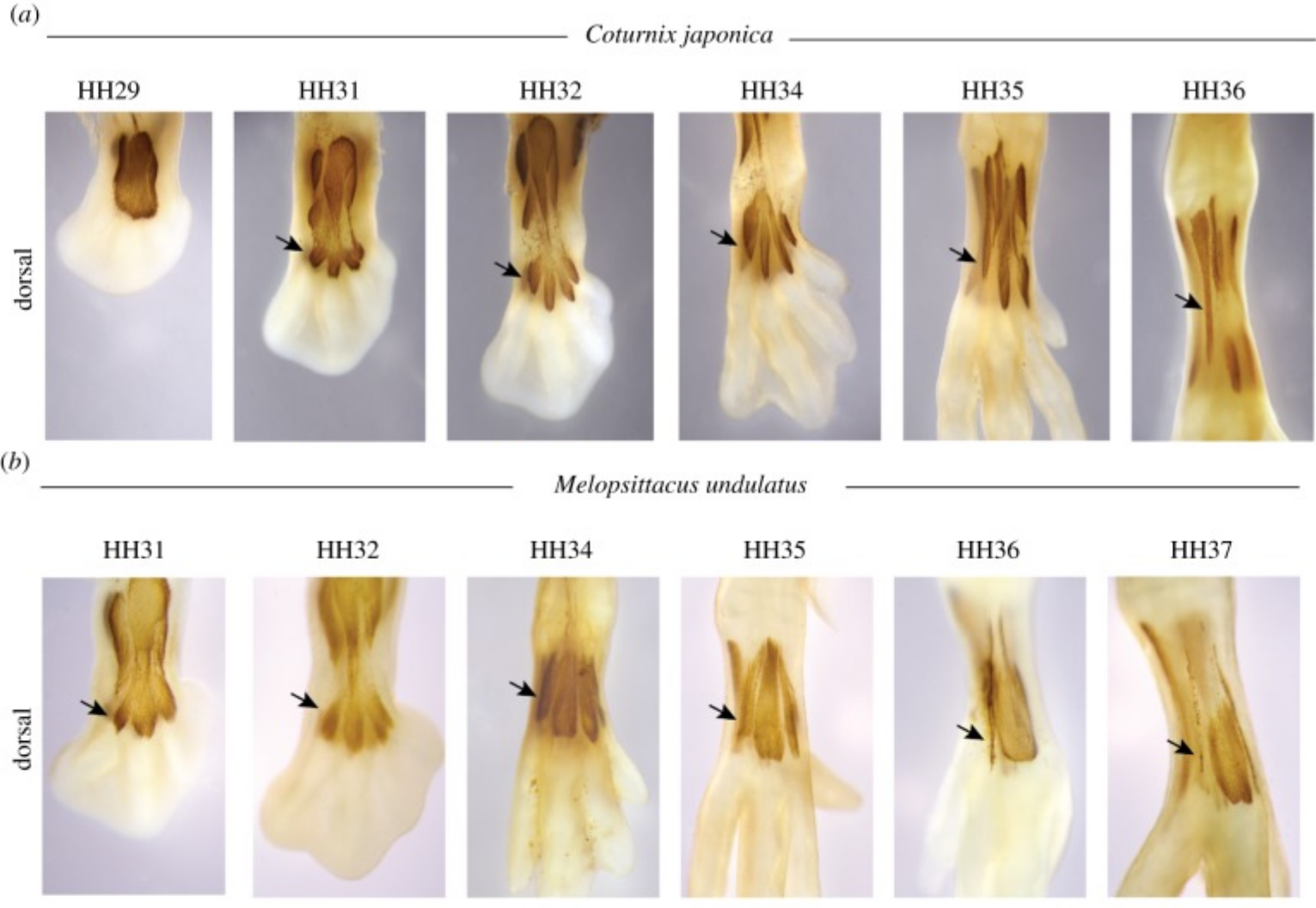 |
Figura 5: Desarrollo del pie de anisodactilo vs cigodáctilo. A. Muestra el desarrollo de un pie de codorniz mientras que B. Muestra el desarrollo de un pie periquito. Brown es la tinción de anticuerpos contra la miosina 2, la cual se encuentra en los músculos. Las flechas señalan el extensor musculus brevis digiti 4, que degenera en el periquito y permite que el músculo abductor digiti 4 tire del cuarto dígito medio-lateralmente. Figura de Botelho et al, 2014. Usado con permiso. |
Hasta donde sabemos, no se ha estudiado el desarrollo de otras confirmaciones de dígitos de las extremidades posteriores de aves. Con base en lo que sabemos sobre el desarrollo del pie de anisodáctilo y cigodáctilo, planteamos la hipótesis de una trayectoria evolutiva y de desarrollo similar para los pies de heterodáctilo. El pie de Heterodáctilo se ve similar al pie de Zygodáctilo pero el dígito 2 se gira en lugar del dígito 4. Pensamos que el desarrollo podría ser el mismo que el del pie Zygodactyl pero los cambios musculares afectan al 2do dígito. Las aves trogonales tienen pies de heterodáctilo (Figuras 1 y 2). Tienen dos dígitos anteriores, los dígitos 3 y 4, y dos posteriores, los dígitos 1 y 2.
Los pies de sindactilo, como los de los martín pescadores (Figura 1) tienen fusión de dos dígitos vía cincha. Debajo de esta cincha, tienen la misma conformación de dígitos que los pies de anisodáctilo. La investigación sobre alas de murciélago ha demostrado que la cincha entre los dígitos se debe a la expresión de un antagonista de BMP. En otros animales como los ratones, los Bmp provocan la muerte del mesénquima interdigital. Entonces, un antagonista de BMP en aves sindactyl podría prevenir potencialmente la apoptosis del mesénquima que rodea los dígitos, conduciendo a pies de sindactilo.
Referencias
Botelho, J.F., Smith-Paredes, D. & Vargas, A.O. (2015) “La altricialidad y la evolución de la orientación del dedo del pie en las aves” Evol Bio l 42:502. https://doi.org/10.1007/s11692-015-9334-7
Alexander O. Vargas, Tiana Kohlsdorf, John F. Fallon, John VandenBrooks, Günter P. Wagner (2008) “La evolución de la expresión de HoxD-11 en el ala de pájaro: Insights from Alligator mississippiensis” PLoS ONE 3 (10): e3325. https://doi.org/10.1371/journal.pone.0003325
João Francisco Botelho, Daniel Smith-Paredes, Daniel Núñez-León, Sergio Soto-Acuña, Alexander O. Vargas (2014) “El origen evolutivo de los pies de cigodáctilo y su posible pérdida en la evolución de Passeriformes” Proc Biol Sci 281 (1788) :20140765. doi: 10.1098/rspb.2014.0765
Scott D. Weatherbee, Richard R. Behringer, John J. Rasweiler, Lee A.Niswander (2006) “La retención de cinchas interdigitales en alas de murciélago ilustra los cambios genéticos subyacentes a la diversificación de las extremidades amniotas” Actas de la Academia Nacional de Ciencias 103 (41) 15103-15107; DOI:10.1073/PNAS.0604934103