
3.23: Difusión, Transporte Activo y Canales de Membrana
- Page ID
- 56601

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
Transporte a través de membranas celulares
Todas las células adquieren las moléculas e iones que necesitan de su fluido extracelular circundante (ECF). Hay un tráfico incesante de moléculas e iones dentro y fuera de la célula a través de su membrana plasmática (Ejemplos: glucosa,\(Na^+\),\(Ca^{2+}\)). En las células eucariotas, también hay transporte dentro y fuera de los compartimentos intracelulares unidos a la membrana, como el núcleo, el retículo endoplásmico y las mitocondrias (Ejemplos: proteínas\(Ca^{2+}\), ARNm y ATP).
Los siguientes problemas pueden ocurrir durante el transporte:
1. Concentraciones relativas
Las moléculas y los iones se mueven espontáneamente por su gradiente de concentración (es decir, de una región de mayor a una región de menor concentración) por difusión. Las moléculas y los iones pueden moverse contra su gradiente de concentración, pero este proceso, llamado transporte activo, requiere el gasto de energía (generalmente del ATP).
2. Las bicapas lipídicas son impermeables a la mayoría de las moléculas e iones esenciales.
La bicapa lipídica es permeable a las moléculas de agua y a algunas otras moléculas pequeñas, sin carga, como el oxígeno (O 2) y el dióxido de carbono (CO 2). Éstas se difunden libremente dentro y fuera de la célula. La difusión del agua a través de la membrana plasmática es de tal importancia para la célula que se le da un nombre especial: ósmosis. Las bicapas lipídicas no son permeables a iones como K + , Na +, Ca 2+ (llamados cationes porque cuando se someten a un campo eléctrico migran hacia el cátodo [la carga negativa electrodo]) y Cl -, HCO 3 - (llamados aniones porque migran hacia el ánodo [el electrodo cargado positivamente]). Tampoco son permeables a pequeñas moléculas hidrófilas como la glucosa y macromoléculas como proteínas y ARN. Las células resuelven el problema del transporte de iones y moléculas pequeñas a través de sus membranas con la ayuda de los siguientes dos mecanismos:
- Difusión facilitada: Las proteínas transmembrana crean un poro lleno de agua a través del cual los iones y algunas pequeñas moléculas hidrófilas pueden pasar por difusión. Los canales se pueden abrir (o cerrar) según las necesidades de la celda.
- Transporte activo: Las proteínas transmembrana, llamadas transportadores, utilizan la energía del ATP para forzar iones o pequeñas moléculas a través de la membrana contra su gradiente de concentración.
Difusión facilitada de iones
La difusión facilitada de iones tiene lugar a través de proteínas, o ensamblajes de proteínas, incrustados en la membrana plasmática. Estas proteínas transmembrana forman un canal lleno de agua a través del cual el ion puede pasar por su gradiente de concentración. Los canales transmembrana que permiten la difusión facilitada pueden ser abiertos o cerrados. Se dice que son “cerrados”; algunos tipos de canales iónicos cerrados:
- ligando-cerrado
- Mecánicamente-cerrado
- voltaje-gated
- luz-cerrado
Canales iónicos con cierre de ligando
Muchos canales iónicos se abren o cierran en respuesta a la unión de una pequeña molécula de señalización o “ligando”. Algunos canales iónicos están regulados por ligandos extracelulares; algunos por ligandos intracelulares. En ambos casos, el ligando no es la sustancia que se transporta cuando se abre el canal.
Ligandos externos
Los ligandos externos (mostrados aquí en verde) se unen a un sitio en el lado extracelular del canal.

Ejemplos:
- Acetilcolina (ACh). La unión del neurotransmisor acetilcolina a ciertas sinapsis abre canales que admiten Na + e inician un impulso nervioso o contracción muscular.
- Ácido gamma amino butírico (GABA). La unión de GABA en ciertas sinapsis, designadas GABA A, en el sistema nervioso central admite iones Cl en la célula e inhibe la creación de un impulso nervioso
Ligandos internos
Los ligandos internos se unen a un sitio en la proteína del canal expuesta al citosol. Ejemplos:
- Los “segundos mensajeros”, como AMP cíclico (AMPc) y GMP cíclico (cGMP), regulan los canales involucrados en la iniciación de impulsos en neuronas que responden a olores y luz respectivamente.
- El ATP es necesario para abrir el canal que permite que los iones cloruro (Cl -) y bicarbonato (HCO 3 -) salgan de la célula. Este canal es defectuoso en pacientes con fibrosis quística. Aunque la energía liberada por la hidrólisis del ATP es necesaria para abrir el canal, este no es un ejemplo de transporte activo; los iones se difunden a través del canal abierto siguiendo su gradiente de concentración.
Canales iónicos con cierre mecánico
Las ondas sonoras que doblan las proyecciones ciliares en las células ciliadas del oído interno abren canales iónicos que conducen a la creación de impulsos nerviosos que el cerebro interpreta como sonido. La deformación mecánica de las células de los receptores de estiramiento abre canales iónicos que conducen a la creación de impulsos nerviosos.
Canales iónicos con regulación de voltaje
En las llamadas células “excitables” como las neuronas y las células musculares, algunos canales se abren o cierran en respuesta a cambios en la carga (medidos en voltios) a través de la membrana plasmática. Por ejemplo, a medida que un impulso pasa por una neurona, la reducción en el voltaje abre canales de sodio en la porción adyacente de la membrana. Esto permite la afluencia de\(Na^+\) dentro de la neurona y con ello la continuación del impulso nervioso. Unos 7000 iones de sodio pasan por cada canal durante el breve periodo (aproximadamente 1 milisegundo) que permanece abierto. Esto se aprendió mediante el uso de la técnica de pinza de parche.
La técnica de la abrazadera de parche
Las propiedades de los canales iónicos se pueden estudiar mediante la técnica de patch clamp. Se presiona una pipeta muy fina (con una abertura de aproximadamente 0.5 µm) contra la membrana plasmática de una célula intacta o la membrana plasmática se puede extraer de la célula y la preparación se coloca en una solución de prueba de la composición deseada. Entonces se puede medir el flujo de corriente a través de un solo canal iónico.

Tales mediciones revelan que cada canal está completamente abierto o completamente cerrado; es decir, la difusión facilitada a través de un solo canal es “todo o ninguno”. Esta técnica ha aportado tanta información valiosa sobre los canales iónicos que sus inventores, Erwin Neher y Bert Sakmann, fueron galardonados con el Premio Nobel en 1991.
Difusión facilitada de moléculas
Algunas moléculas orgánicas hidrófilas pequeñas, como los azúcares, pueden pasar a través de las membranas celulares mediante difusión facilitada. Una vez más, el proceso requiere proteínas transmembrana. En algunos casos, estos —como los canales iónicos— forman poros llenos de agua que permiten que la molécula pase dentro (o fuera) de la membrana siguiendo su gradiente de concentración.
Ejemplo:
Maltoporina. Este homotrímero en la membrana externa de E. coli forma poros que permiten que el disacárido maltosa y algunas moléculas relacionadas se difundan en la célula.
Ejemplo: La membrana plasmática de glóbulos rojos humanos contiene proteínas transmembrana que permiten la difusión de glucosa desde la sangre hacia la célula.
Obsérvese que en todos los casos de difusión facilitada a través de canales, los canales son selectivos; es decir, la estructura de la proteína admite solo ciertos tipos de moléculas a través. Aún no se ha probado si todos los casos de difusión facilitada de moléculas pequeñas utilizan canales. Quizás algunas moléculas pasan a través de la membrana por un cambio conformacional en la forma de la proteína transmembrana cuando se une a la molécula a transportar.
En cualquier caso, la interacción entre la molécula que se transporta y su transportador se asemeja en muchos sentidos a la interacción entre una enzima y su sustrato.
Transporte Activo
El transporte activo es el bombeo de moléculas o iones a través de una membrana contra su gradiente de concentración. Requiere de una proteína transmembrana (generalmente un complejo de ellas) llamada transportador y energía. La fuente de esta energía es el ATP.
La energía del ATP puede ser utilizada directa o indirectamente.
- Transporte Activo Directo. Algunos transportadores se unen directamente al ATP y utilizan la energía de su hidrólisis para impulsar el transporte activo.
- Transporte Activo Indirecto. Otros transportadores utilizan la energía ya almacenada en el gradiente de un ion directamente bombeado. El transporte activo directo del ion establece un gradiente de concentración. Cuando esto es aliviado por la difusión facilitada, la energía liberada puede ser aprovechada para el bombeo de algún otro ion o molécula.
Transporte Activo Directo
La Na + /K + ATPasa
El citosol de las células animales contiene una concentración de iones potasio (K +) hasta 20 veces mayor que la del líquido extracelular. Por el contrario, el fluido extracelular contiene una concentración de iones de sodio (Na +) hasta 10 veces mayor que la del interior de la célula. Estos gradientes de concentración se establecen por el transporte activo de ambos iones. Y, de hecho, el mismo transportador, llamado Na + /K + ATPasa, hace ambos trabajos. Utiliza la energía de la hidrólisis de ATP para
- transportar activamente 3 iones Na + fuera de la célula
- por cada 2 K + iones bombeados a la celda.
Esto logra varias funciones vitales:
- Ayuda a establecer una carga neta a través de la membrana plasmática con el interior de la célula cargado negativamente con respecto al exterior. Este potencial de reposo prepara a las células nerviosas y musculares para la propagación de potenciales de acción que conducen a impulsos nerviosos y contracción muscular.
- La acumulación de iones de sodio fuera de la célula extrae el agua de la célula y así le permite mantener el equilibrio osmótico (de lo contrario se hincharía y estallaría desde la difusión interna del agua).
- El gradiente de iones de sodio se aprovecha para proporcionar la energía para ejecutar varios tipos de bombas indirectas.
Los papeles cruciales de la ATPasa Na + /K + se reflejan en el hecho de que casi un tercio de toda la energía generada por las mitocondrias en las células animales se usa solo para hacer funcionar esta bomba.
La ATPasa H + /K +
Las células parietales de tu estómago utilizan esta bomba para secretar jugo gástrico. Estas células transportan protones (H +) desde una concentración de aproximadamente 4 x 10 -8 M dentro de la célula hasta una concentración de aproximadamente 0.15 M en el jugo gástrico (dándole un pH cercano a 1). No es de extrañar que las células parietales estén rellenas de mitocondrias y utilicen enormes cantidades de ATP ya que llevan a cabo esta concentración de protones de tres millones de veces.
Las ATPasas Ca 2+
Una ATPasa Ca 2+ se localiza en la membrana plasmática de todas las células eucariotas. Utiliza la energía proporcionada por una molécula de ATP para bombear un ion Ca 2+ fuera de la célula. La actividad de estas bombas ayuda a mantener el gradiente de concentración de ~20.000 veces de Ca 2+ entre el citosol (~ 100 nM) y la ECF (~ 20 mM). En el músculo esquelético en reposo, hay una concentración mucho mayor de iones calcio (Ca 2+) en el retículo sarcoplásmico que en el citosol. La activación de la fibra muscular permite que parte de este Ca 2+ pase por difusión facilitada al citosol donde desencadena la contracción.
Después de la contracción, este Ca 2+ se bombea de nuevo al retículo sarcoplásmico. Esto lo hace otra ATPasa Ca 2+ que utiliza la energía de cada molécula de ATP para bombear 2 iones Ca 2+.
Bombas 1. - 3. se designan transportadores de iones tipo P porque utilizan el mismo mecanismo básico: un cambio conformacional en las proteínas ya que son fosforiladas reversiblemente por ATP. Y las tres bombas se pueden hacer para funcionar hacia atrás. Es decir, si se permite que los iones bombeados se difundan de nuevo a través del complejo de membrana, el ATP se puede sintetizar a partir de ADP y fosfato inorgánico.
Transportadores ABC
Los transportadores ABC (” A TP- B inding C assette”) son proteínas transmembrana que
- exponer un dominio de unión a ligando en una superficie y
- Dominio de unión a ATP en la otra superficie.
El dominio de unión al ligando generalmente se restringe a un solo tipo de molécula.
El ATP unido a su dominio proporciona la energía para bombear el ligando a través de la membrana.
El genoma humano contiene 48 genes para transportadores ABC. Algunos ejemplos:
- CFTR: el regulador de la conductancia transmembrana de la fibrosis quística
- TAP, el transportador asociado al procesamiento de antígenos
- El transportador que utilizan las células hepáticas para bombear las sales de los ácidos biliares hacia la bilis.
- Transportadores ABC que bombean fármacos quimioterapéuticos fuera de las células cancerosas, reduciendo así su efectividad.
Los transportistas ABC deben haber evolucionado temprano en la historia de la vida. Los dominios de unión a ATP en arqueas, eubacterias y eucariotas comparten una estructura homóloga, el “casete” de unión a ATP.
Transporte Activo Indirecto
El transporte activo indirecto utiliza el flujo descendente de un ion para bombear alguna otra molécula o ion contra su gradiente. El ion conductor suele ser sodio (Na +) con su gradiente establecido por la ATPasa Na + /K +.
Bombas Symport
En este tipo de transporte activo indirecto, el ion impulsor (Na +) y la molécula bombeada pasan a través de la bomba de membrana en la misma dirección. Ejemplos:
- El transportador de Na + /glucosa. Esta proteína transmembrana permite que los iones sodio y la glucosa ingresen juntos a la célula. Los iones de sodio fluyen por su gradiente de concentración mientras que las moléculas de glucosa son bombeadas por el suyo. Posteriormente el sodio es bombeado de nuevo fuera de la celda por la ATPasa Na + /K +. El transportador de Na + /glucosa se utiliza para transportar activamente la glucosa fuera del intestino y también fuera de los túbulos renales y de regreso a la sangre.
- Todos los aminoácidos pueden ser transportados activamente, por ejemplo fuera de los túbulos renales y hacia la sangre, mediante bombas symport impulsadas por sodio.
- Las bombas symport impulsadas por sodio también devuelven los neurotransmisores a la neurona presináptica.
- El transportador de Na + /yoduro. Este simporter bombea iones yoduro a las células de la glándula tiroides (para la fabricación de tiroxina) y también a las células de la glándula mamaria (para suplir la necesidad de yoduro del bebé).
- La permeasa codificada por el operón lac de E. coli que transporta lactosa a la célula.
Bombas Antiport
En las bombas antipuerto, el ion impulsor (nuevamente, generalmente el sodio) se difunde a través de la bomba en una dirección proporcionando la energía para el transporte activo de alguna otra molécula o ion en la dirección opuesta. Ejemplo:
Los iones Ca 2+ son bombeados fuera de las células mediante bombas antipuerto impulsadas por sodio. Las bombas antiportuarias en la vacuola de algunas plantas aprovechan la difusión facilitada hacia afuera de protones (ellos mismos bombeados en la vacuola por una H+ ATPasa) para el transporte activo hacia adentro de iones de sodio. Esta bomba antipuerto de sodio/protón permite a la planta secuestrar iones de sodio en su vacuola. Las plantas transgénicas de tomate que sobreexpresan esta bomba antiportuaria de sodio/protón son capaces de prosperar en suelos salinos demasiado salados para los tomates convencionales. Bombas antiportuarias para el transporte activo hacia el interior de iones nitrato (NO 3 −)
Algunas enfermedades hereditarias de los canales iónicos
Se ha descubierto que un número creciente de enfermedades humanas son causadas por mutaciones heredadas en genes que codifican canales.
Ejemplos:
- Enfermedades de los canales de cloruro
- fibrosis quística
- tendencia hereditaria a cálculos renales (causada por un tipo de canal de cloruro diferente al involucrado en la fibrosis quística)
- Enfermedades de los canales de potasio
- la mayoría de los casos de síndrome de QT largo, un trastorno hereditario del latido del corazón
- una rara tendencia hereditaria a las crisis epilépticas en el recién nacido
- varios tipos de sordera heredada
- Enfermedades de los canales de sodio
- tendencia hereditaria a ciertos tipos de espasmos musculares
- Síndrome de Liddle. El transporte inadecuado de sodio fuera de los riñones, debido a un canal de sodio mutante, conduce a una presión osmótica elevada de la sangre y la consiguiente hipertensión (presión arterial alta)
Osmosis
La ósmosis es un término especial utilizado para la difusión del agua a través de las membranas celulares. Aunque el agua es una molécula polar, es capaz de pasar a través de la bicapa lipídica de la membrana plasmática. Las acuaporinas —proteínas transmembrana que forman canales hidrofílicos— aceleran en gran medida el proceso, pero incluso sin éstas, el agua todavía es capaz de atravesar. El agua pasa por difusión de una región de mayor a una región de menor concentración. Tenga en cuenta que esto se refiere a la concentración de agua, NO a la concentración de ningún soluto presente en el agua. El agua nunca se transporta activamente; es decir, nunca se mueve contra su gradiente de concentración. Sin embargo, la concentración de agua puede ser alterada por el transporte activo de solutos y de esta manera se puede controlar el movimiento del agua dentro y fuera de la celda. Ejemplo: la reabsorción de agua de los túbulos renales de vuelta a la sangre depende del agua que siga detrás del transporte activo de\(Na^+\).
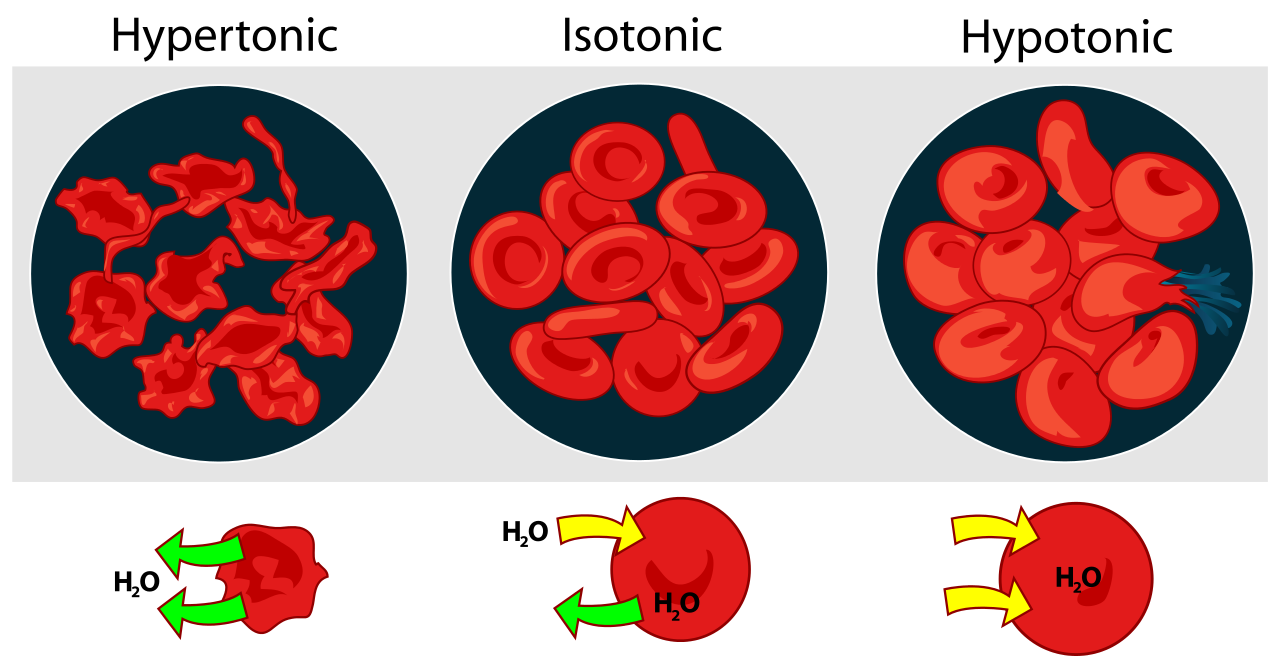
- Soluciones hipotónicas: Si la concentración de agua en el medio que rodea a una célula es mayor que la del citosol, se dice que el medio es hipotónico. El agua ingresa a la célula por ósmosis. Un glóbulo rojo colocado en una solución hipotónica (e.g., agua pura) estalla inmediatamente (“hemólisis”) por la afluencia de agua. Las células vegetales y las células bacterianas evitan estallar en entornos hipotónicos por sus fuertes paredes celulares. Estos permiten la acumulación de turgencia dentro de la celda. Cuando la presión de turgencia es igual a la presión osmótica, cesa la ósmosis.
- Soluciones isotónicas: Cuando los glóbulos rojos se colocan en una solución salina al 0.9%, no ganan ni pierden agua por ósmosis. Se dice que tal solución es isotónica. El líquido extracelular (ECF) de las células de mamíferos es isotónico para su citoplasma. Este equilibrio debe mantenerse activamente debido al gran número de moléculas orgánicas disueltas en el citosol pero no presentes en la ECF. Estas moléculas orgánicas ejercen un efecto osmótico que, de no compensarse, provocaría que la célula absorbiera tanta agua que se hincharía e incluso podría estallar. Este destino se evita bombeando iones de sodio fuera de la célula con la ATPasa Na + /K +.
- Soluciones hipertónicas: Si los glóbulos rojos se colocan en agua de mar (aproximadamente 3% de sal), pierden agua por ósmosis y las células se marchitan. El agua de mar es hipertónica para su citosol. De manera similar, si se coloca un tejido vegetal en agua de mar, el contenido celular se encoge lejos de la pared celular rígida. Esto se llama plasmólisis. El agua de mar también es hipertónica para la ECF de la mayoría de los vertebrados marinos. Para evitar la deshidratación fatal, estos animales (por ejemplo, peces óseos como el bacalao) deben beber continuamente agua de mar y luego desaltarla bombeando iones fuera de sus agallas mediante transporte activo.
Las aves marinas, que pueden pasar largos períodos de tiempo lejos del agua dulce, y las tortugas marinas utilizan un dispositivo similar. Ellos, también, beben agua salada para atender sus necesidades de agua y utilizan energía metabólica para desalarla. En la gaviota arenque, aquí se muestra, la sal es extraída por dos glándulas en la cabeza y liberada (en una solución muy concentrada —es más salada que la sangre) hacia el exterior a través de las fosas nasales. Las serpientes marinas utilizan un mecanismo de desalación similar.
